-
真核起始因子 编辑
真核起始因子(英文:eukaryotic initiation factor,简称为eIF),又称为真核翻译起始因子,是指参与真核翻译起始这一过程的蛋白质。与原核起始因子只有三种(IF1、IF2、IF3)相比,真核起始因子种类多且复杂,已鉴定的真核起始因子共有12种。通过这些真核起始因子之间以及不同的真核起始因子与核糖体、mRNA和起始tRNA之间的相互作用,来完成真核生物的翻译起始。因此相比于原核生物,真核生物的翻译起始过程更多得依赖于蛋白质与蛋白质以及蛋白质与RNA之间的相互作用,而非RNA与RNA间的相互作用。
除了参与真核翻译起始进程之外,许多真核起始因子还具有其他一些功能。例如,eIF3参与细胞生长和细胞周期的调控。
多个真核起始因子也与疾病相关。例如,eIF2B的两个不同亚基的编码基因的突变能够导致一种被称为“白质消失”(vanishing white matter)的严重的遗传性神经退行性疾病;eIF3的e亚基与癌症密切相关。

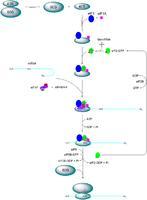
真核起始因子之间与核糖体之间存在着大量的相互作用,构成了一个相互作用网络。其中,eIF3是介导这一相互作用网络的中心点,它的一个或多个亚基可以与eIF1、eIF1A、eIF2、eIF4B、eIF4G、eIF5以及核糖体40S亚基相互作用。这些相互作用可能是稳定的,从而可以形成稳定的复合物参与翻译起始进程;同样也可能有许多动态的相互作用,使得翻译起始进程是在可调控的情况下进行。
稳定相互作用
许多研究结果显示,eIF1、eIF3、eIF2·GTP·Met-tRNAi和eIF5能够在酵母体内结合,形成稳定的多因子复合物(MFC),而且MFC的完整性对于翻译起始,特别是在AUG密码子的选择中十分重要。而MFC是以eIF5作为核心,通过eIF5的C端结构域与eIF1、eIF2的β亚基以及eIF3的c亚基相互作用来聚合形成的。
动态相互作用
真核起始因子之间的相互作用很多可能是动态的,而且只发生在起始进程中的特定点上。有多个实验现象都支持这一观点:
(1) 在一些因子与40S亚基解离后,另一些因子就可以与40S亚基结合;
(2) 发现在细胞中,形成复合物的一些因子的浓度要高于另一些因子,这就表明这些高浓度的因子部分参与形成复合物,而另一部分则不参与;
(3) 在一些例子中,两个因子间的相互作用对于第三个因子能产生排斥,提示它们只能在起始进程的不同时刻发挥作用。例如,eIF1和eIF5的相互作用与eIF4G彼此排斥,eIF2和eIF4G的相互作用与eIF5,以及eIF5和eIF2B的相互作用与eIF2也相互排斥。
已知的真核起始因子中,eIF2B与人类遗传病的关系最为密切。eIF2B的五个亚基基因的常染色体遗传性隐性突变会导致白质异常,在临床上表现为一系列严重的连续症状,称为“eIF2B相关紊乱”。典型的如脑白质病,即白质消失(vanishing white matter,VWM)和卵巢衰竭(ovarian failure)。这种紊乱疾病持续时间久,伴随年龄增长而不断恶化,而且当感染发烧或轻微脑部外伤都可以恶化病情而导致死亡。最严重时会在婴儿期就造成死亡,而如果持续到最后会导致成人卵巢发育失败并可能伴随神经退化。
在哺乳动物中,eIF3的e亚基(eIF3e)由Int6基因编码,而Int6基因是鼠乳腺癌病毒(mouse mammary tumor virus)基因组的整合位点。而病毒基因组的插入能够导致生成被剪切的eIF3e,表达被剪切的eIF3e就能够导致细胞发生癌变。因此,eIF3e虽然不直接引发癌症,但却与癌症的发生密切相关。
此外,eIF2的α亚基中磷酸化位点的突变能够导致与PERK(PKR-like endoplasmic reticulum kinase)基因缺失相似的症状。而PERK基因突变能够遗传性疾病Wolcott-Rallison综合症,表现为幼年重糖尿病伴随骨骼异常生长延迟。但还没有关于该磷酸化位点的突变发生在人体中的报道。