-
β氧化 编辑
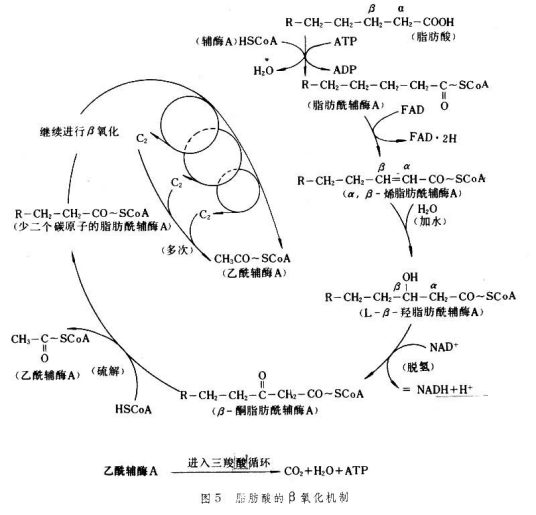
定义:脂肪酸在一系列酶的作用下,在α碳原子和β碳原子之间断裂,生成乙酰辅酶A,和较原来少两个碳原子的脂肪酰辅酶A。
在肝脏内脂肪酸经β-氧化作用生成乙酰辅酶A,两分子的乙酰辅酶A可缩合生成乙酰乙酸。乙酰乙酸可脱羧生成丙酮,也可还原生成β-羟丁酸。乙酰乙酸、β-羟丁酸和丙酮总称为酮体。肝脏不能利用酮体,必须经血液运至肝外组织特别是肌肉和肾脏,再转变为乙酰辅酶A而被氧化利用。酮体作为有机体代谢的中间产物,在正常的情况下,其产量甚微,患糖尿病或食用高脂肪膳食时,血中酮体含量增高.尿中也能出现酮体 。
脂肪酸是由一条长的烃基上附加一个羧基的化合物,溶解度一般不大,主要来源于脂肪在人体消化道内的水解。
碳原子个数为偶数的脂肪酸进入人体后,其羧基在细胞质基质中与乙酰辅酶A(乙酰CoA)结合,之后循环往复地被催化脱去乙基,产生新的乙酰CoA,直至碳原子全部脱去。
新产生的乙酰CoA大多进入线粒体基质中脱羧脱氢,进而被柠檬酸合成酶催化产生柠檬酸,参与三羧酸循环(又名Kreb循环或柠檬酸循环)。
(1)脂肪酸的活化:脂肪酸的氧化首先须被活化,在ATP、CoA-SH、Mg2+存在下,由位于内质网及线粒体外膜的脂酰CoA合成酶,催化生成脂酰CoA。活化的脂肪酸不仅为一高能化合物,而且水溶性增强,因此提高了代谢活性。
(2)脂酰CoA的转移:是在胞液中进行的,而催化脂肪酸氧化的酶系又存在于线粒体基质内,故活化的脂酰CoA必须先进入线粒体才能氧化,但已知长链脂酰辅酶A是不能直接透过线粒体内膜的,因此活化的脂酰CoA要借助肉碱(camitine),即L-3羟-4-三甲基铵丁酸,而被转运入线粒体内,在线粒体内膜的外侧及内侧分别有肉碱脂酰转移酶I和酶Ⅱ,两者为同工酶。位于内膜外侧的酶Ⅰ,促进脂酰CoA转化为脂酰肉碱,后者可借助线粒体内膜上的转位酶(或载体),转运到内膜内侧,然后,在酶Ⅱ催化下脂酰肉碱释放肉碱,后又转变为脂酰CoA。这样原本位于胞液的脂酰CoA穿过线粒体内膜进入基质而被氧化分解。一般10个碳原子以下的活化脂肪酸不需经此途径转运,而直接通过线粒体内膜进行氧化。
 β-氧化途径
β-氧化途径
a 脱氢:脂酰CoA在脂酰基CoA脱氢酶的催化下,其烃链的α、β位碳上各脱去一个氢原子,生成α、β烯脂酰CoA(trans-y-enoylCoA),脱下的两个氢原子由该酶的辅酶FAD接受生成FADH2。后者经电子传递链传递给氧而生成水,同时伴有1.5分子ATP的生成。
b 加水:α、β烯脂酰CoA在烯酰CoA水合酶的催化下,加水生成β-羟脂酰CoA(βhydroxyacylCoA)。
c 再脱氢:β-羟脂酰CoA在β-羟脂酰CoA脱氢酶(L-βhydroxyacylCoAdehydrogenase)催化下,脱去β碳上的2个氢原子生成β-酮脂酰CoA,脱下的氢由该酶的辅酶NAD+接受,生成NADH+H+。后者经电子传递链氧化生成水及2.5分子ATP。
d 硫解:β-酮脂酰CoA在β-酮脂酰CoA硫解酶(β-ketoacylCoAthiolase)催化下,加一分子CoASH使碳链断裂,产生乙酰CoA和一个比原来少两个碳原子的脂酰CoA。以上4步反应均可逆行,但全过程趋向分解,尚无明确的调控位点。
1分子软脂酸含16个碳原子,靠7次β氧化生成7分子NADH+H+,7分子FADH2,8分子乙酰CoA,共生成:7×1.5+7×2.5+8×10=108分子ATP,而所有脂肪酸活化均需耗去2分子ATP。故1分子软脂酸彻底氧化净生成:7×1.5+7×2.5+8×10-2=106分子ATP。
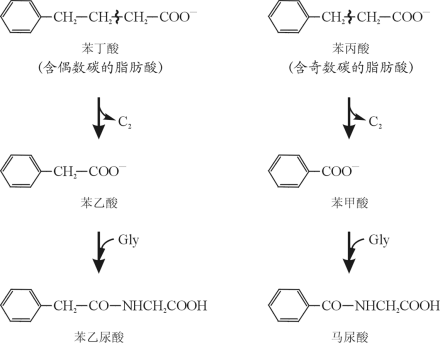
以后的实验证明Knoop推测的准确性,由此提出了脂肪酸的β-氧化作用。
β-氧化作用是指脂肪酸在β-碳原子上进行氧化,然后α-碳原子和β-碳原子之间键发生断裂。每进行一次β-氧化作用,分解出一个二碳片段,生成较原来少两个碳原子的脂肪酸。
后来对CoA的发现以及分离和提纯了参与脂肪酸氧化的各种酶,更弄清了其氧化机制的细节。E.P.Kennedy 和 A.L.Lehninger(1949)指出此氧化系统存在于线粒体中,后来D.E.Green及F.Lynen(1953)各自独立地从线粒体的丙酮粉末提取出可溶性酶,成功地分离出β氧化各个阶段的酶,明确了脂肪β氧化,按下述过程进行:
(1)由脂肪酸活化酶使脂肪酸与 CoA结合,
(2)由乙酰CoA脱氢酶的作用使乙酰CoA脱氢,
(3)由烯酰CoA水合酶的作用使烯酰CoA加水,
(4)由β-羟基乙酰 CoA脱氢酶的作用使β-羟基乙酰 CoA脱氢,
(5)由β-酮酰CoA硫解酶的作用使β酮酰CoA裂解。
经以上5个阶段逐次游离出来的乙酰CoA(C2片段)经三羧酸循环而氧化。其能量收支为每分子棕榈酸(C16)产生130分子ATP。不饱和脂肪酸的氧化除需上述各种酶之外,还需要催化3-顺-烯酰CoA转变成2-反式的3-顺, 2-反-烯酰CoA异构酶和催化D(一)-3-羟式成L(+)-3-羟式的3-羟乙酰CoA-3-表异构酶参与。由奇数C原子脂肪酸分解产生的丙酰CoA,通过羧化及异构化而转变成琥珀酰CoA再进一步变化 。