-
转座因子 编辑
中文名:转座因子
外文名:transposableelement
细胞中能改变自身位置的一段脱氧核糖核酸(DNA)序列。转座因子改变位置(例如从染色体上的一个位置转移到另一个位置,或者从质粒转移到染色体上)的行为称为转座(transposition)。
由于转座因子既能给基因组带来新的遗传物质,在某些情况中又能像一个开关那样启动或关闭某些基因,并常使基因组发生缺失、重复或倒位等DNA重排,所以它与生物演化有密切的关系,并可能与个体发育、细胞分化有关。
此外,带有不同抗药性基因的转座因子在细菌质粒间的转座会导致多价抗药性质粒的形成,这将使多种药物药效降低,对人类的健康是一个威胁,因此需要研究解决的办法。另一方面具有独特功能的转座因子已经成为遗传学研究中一种有用的工具。
在50年代以前,人们对于基因的认识一般是每一个基因组的DNA的量是固定的,它包括数目固定,位置固定、功能固定的一系列基因,以保持生物性状能稳定地遗传下去。但同时,基因也会发生突变。一般自发突变的频率是很低的,当然也存在着高突变频率的现象,这说明在基因组中存在高度不稳定的基因,很长时间人们忽视了这种现象。第一个注意到不稳定基因的人是De.Vires,他在19世纪末就研究了金鱼草花的颜色的不稳定突变,但没能作出理论解释。20世纪的前几十年内,R.A.Emesor研究了能干扰种皮中桔红色素合成的玉米突变品系,他提出籽粒发育过程中发生回复突变,形成有色细胞和无色细胞交替的条纹是由不稳定的隐性基因造成的,但他不知道是什么原因使隐性城因不稳定。虽然McClintock早在1951年就在玉米中发现了转座因子,但由于人们受到基因在染色体上有序排列的传统观念的桎梏而难以接受“跳跃基因”这一新的概念,所以直到1967年Shapiro才在E.coli中发现了转座因子(transposable element)。他在半乳糖操纵子中发现了一种极性突变。gal操纵子有三个基因:gal K,T,E,它们编码三种酶催化Gal的分解代谢。其中间产物Gal-1-p是有毒的,因此galT-在含半糖培养基上因使Gal-1-p积累而难以成活。那么在此培养基上能生长的各种突变型,应是galt-的回复突变及galK-galT-(因它不能产生Gal-1-p),但意外发的发现galE-galT-也能生存,按理说这种突变型可使Gal-1-p积累是不可能存活的,但却居然生长良好,其原因何在呢?经进一步研究发现galE-是极性突变,由于它的突变使远离操纵位点的galK活性大大下降,因而细胞中不会积累Gal-1-p之故,从中首次分离了转座因子。分析转座因子的物理特性,更证实了转座因子理论的正确。已从好几种原核和真核生物中分离到转座因子,并在进行DNA水平的研究 。
自从1928年,F.Criiffith所作的用肺突球菌对小家鼠的感染实验,弄清了导致由R菌株转化成S菌株的转化因子就是DNA后,直到1943年才确定细菌突变的自发性的本质。以后大量的实验证明,在细菌中转移的传导现象的产生是由于菌体内有一种活动因子——转座因子的存在。下面就其原核生物的转座因子的类别和遗传学效应的研究结果作一概述 。
原核生物转座因子的类别
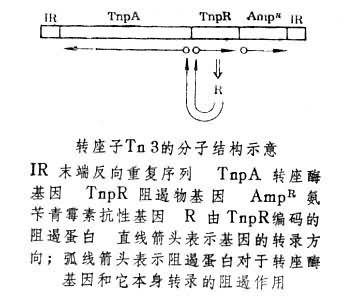
转座因子大致分为三类:(1)插入序列 , 这是一类除了和它的转座作用有关的基因以外不带有任何其他基因的转座因子。它们是较小的转座因子,可以在染色体、质粒上发现它们。F因子和大肠杆菌的染色体上有一些相同的插入序列(IS) ,现已搞清至少有五种DNA插入序列,它既可独立地,也可以作为其他转座因子的一个部分存在于某些细菌的染色体和质粒上。(2)转座子,以Tn表示,首先在质体中发现了这种转座子。转座子的分子两端有相同的序列,如果它们的方向相反,则称为反向重复顺序IR,某些转座子的IR便是已知的侣因子,这些IR既可以作为Tn的一个部分而转座,也可以单独转座。转座子除含有与转座有关的基因和核昔酸序列外,还含有一些其它的基因。(3)转座噬菌体、温和噬菌体既能以自主的自我复制颗粒而存在,也可以插入延续的细菌染色体而与其一起复制,宛如附加的细菌基因群。最著名的温和噬菌体是入,久也浸染大肠杆菌,它既可以像烈性噬菌体一样裂解寄主细胞释放出几百个成熟的噬菌体,也可以成为原噬菌体和寄主染色体之间呈半永久联合,当细菌染色体复制时入噬菌体也跟着一起复制。1953年发现,在大肠杆菌敏感菌体和溶源性菌株间的F十又F一杂交中,入原噬菌体和细菌gal基因一起分离。1955年证明入原噬菌体和gal可被噬菌体lP共同转导。1956年发现了入对gal的限制性转导。这些研究表明入噬菌体具有转座子的性质。另外大肠杆菌的噬菌体Mu,可以引起肠菌的几乎任何一个基因发生插入突变,所以它也具有转座因子的特性。
原核生物的转座因子的遗传学效应
虽然各种转座因子上所带的基因可以很不相同,但它们都有一些共同的遗传学效应:
(1)引起插入突变,使插入位置上出现新的基因。除了上面已经提到的由Is1和Mu引起的插入突变以外,Tn同样能引起插入突变,那么,不管插入的转座因子上带有何种基因,它一方面造成一个基因的插入突变,另一方面使这一位置上出现了个新的基因。
(2)促使染色体发生畸变。由于Is1的存在,它的侧挥容易发生缺失,缺失发生的频率高出自发缺失频率10-1000倍。
(3)切离。转座因子可以从原来的位置上消失,这一过程称为切离,准确的切离使插入失活的基因发生回复突变,不准确的切离并不带来回复突变,而是带来染色体畸变。
(4)转座因子转移到新的位置上后,原有位置上的转座因子保持不变。
1947年美国冷泉港实验室的“玉米夫人”McClintock首先在玉米中发现并描述了转座因子。转座因子的发现,打破了传统遗传学上关于基因在染色体上固定排列及同源染色体交换的观念,揭示了基因的流动性,具有重要的意义 。
转座因子的结构特点和分类
至少在32种植物上有转座因子存在,其中研究最多的是玉米、金鱼草、拟南芥等。其中玉米的Ac/Ds 转座因子和金鱼草的Tam转座因子是研究和利用最多的植物转座因子。植物转座因子具有特定的序列特点,大小一般在1.4~17kb之间,根据DNA片段的序列可初步确定某段DNA是转座因子。根据基因结构特点,转座因子可分为:简单转座因子和复合转座因子。根据转座机制,可分为两大类:通过DNA中间体进行转座DNA转座因子和通过RNA中间体进行转座的逆转录转座因子。
根据序列特点,也可进一步将DNA 转座因子分为以下几类。(1)Ac/Ds类:该类包括最早确定的转座因子,即玉米中的激活/解离因子(Ac/Ds)。对这类因子及其整合位点的分子分析表明,在Ac和Ds的两端都带有11bp末端反向重复序列,在它们的插入靶位点上都有8bp的正向重复(direCT repeats, DRs)。虽然在多种植物种类中Ac/Ds因子都能作为转基因而起作用,但只在玉米中发现该类因子。(2)CACTA类:这一类转座因子通常较大(可达15.2kb)。其TIRs具保守序列CACTA。可移动的CACTA因子在大豆、金鱼草和玉米等植物中都已发现。(3)MITE类:最近确定的最大的一类植物转座因子为MITE(miniature invERTed repeattransposable elements)类。MITE在大多数植物的基因组中普遍存在,其特征是较大(100~300kb),具有8~12bp的TIRs和在插入位点的一般2~3bp的DRs,MITE优先插入基因的非编码区。迄今,已确定的MITE可被进一步分为七类:Tourist, Stowaway, Gaijin, Castaway, Ditto, Wanderer和Explorer。除了DNA转座因子外,在植物基因组中已确定了大量的逆转录转座因子,即在植物基因组中的大量的分散的重复序列,它们通过RNA中间物实现转位。逆转录转座因子两侧都有长的末端重复序列(long terminalrepeats, LTRs),能编码发生转位作用所需的酶。已发现许多逆转录转座因子包含有与反转录病毒的相类似的结构,如逆转录转座因子都具有逆转录酶或类似反转录病毒的反转录酶的开放阅读框架(ORF)。许多逆转录转座因子的逆转录酶基因含有具内切酶活性的成分,还具有编码反转录病毒蛋白的基因,即结构蛋白编码区(gag)、具有多种酶活性的蛋白编码区(pol)和蛋白酶编码区(pro)。在植物中确定的第一个完整的逆转录转座因子是番茄的Tnt1因子,另外,在玉米、番茄和水稻中都已发现了逆转录转座因子。
转座因子的作用
转座因子作用的两种假说。假说一:作物在不良外界环境的压力下,如组织培养,辐射或病原侵染等,会发生基因重组,而转座因子在这种重组中起重要作用。支持这种假说的证据有:(1)番茄的Tnt1转座因子的转位可由广谱的微生物和真菌的激发子来诱导;(2)在水稻中,组织培养条件下可激发Tos10、Tos17和Tos19的转位,这种转位的激发可引起水稻基因组中这些因子的拷贝数的增加,还可引起多个水稻基因的插入突变;(3)转座因子在调节区和编码区的插入和切除可改变基因的编码能力和表达方式;(4)在玉米的研究中,在Adh1-F和u22位点的间隔区确定了10个逆转录转座因子家族,它们占核DNA的50%。假说二:植物具有一种遗传机制来产生显性的抗病专化性以对抗病原物小种的毒性变化,而抗病基因多样化的一个可能的机制就是转座因子诱发的基因改变。支持这一假说的分子生物学证据有:在Hm1基因中发现了一段315个bp的插入序列(命名为dHBr),它可破坏该基因的抗性作用。在植物的进化中,转座因子也在不断进化,由于重组、缺失等由活性的转座子而丧失转位能力,成为稳定基因组结构中的一部分。同时产生新的有活性的转座子。如对水稻抗病基因Xa21家族7个成员的17个转座因子的序列分析表明,与更早的进化,如复制、重组相比,许多转座因子好像是最近才具活性。
转座因子在分子生物学研究中的应用
随着人们对转座因子的转位机制和作用研究的逐步深入,转座因子的应用也越来越广泛,其中主要有以下三个方面的应用。(1)遗传育种上的应用。一方面,转座子的转位会在靶位点引起其邻近基因序列和功能的变化而引起突变。因此,可根据转座子转座的遗传学效应来筛选突变体,培育新品种。另一方面,某些转座子可能是调节基因活动的开关。如玉米子粒各种花斑类型是Ac和Ds等多个基因相互作用的结果。其中Ac可起调节因子的作用,Ds则起调节基因的作用。因此转座子的研究对于培育作物具重要意义。此外,转座子可作为基因定位的标记而用于基因的定位、分离和克隆。(2)转座子标签技术克隆目的基因。利用转座子作为标签分离和克隆外源目的基因的基本原理为:转座子或T-DNA形式的转座子转座插入到靶位点会引起靶位点基因或邻近基因的突变,转座子插入到目的基因后利用转座子上的标记基因就可检测目的基因的位置,利用转座子序列设计探针则可克隆目的基因。该技术已得到成功的应用,用该技术已克隆的基因有玉米的Hm1基因,番茄的cf-9基因,烟草的N基因和亚麻的L6基因等。可利用的植物转座子系统有玉米的Ac/Ds转座子系统和Enchancer(即Spm)系统及金鱼草的Tam3系统等。(3)转座子载体系统介导基因转化。由于转座子可在植物基因组中自主地发生转座而插入到新位点,因此,把目的基因与转座子构建成新的质粒就可通过农杆菌介导等方法将目的基因转化到植物基因组中。转座子介导基因转化的优点是:转座子的流动性的特点可有利于目的基因的插入和整合;同时利用转座子的某些内切酶的作用可消除转基因植株中的选择标记基因,从而确保转基因产品的安全性。另外,近年来在真核生物中发现的逆转录转座子,由于它们在转位过程中需要以RNA为中间体经逆转录再分散到基因组中,这些动态的DNA片段将在基因工程中作为理想的目的基因载体系统。