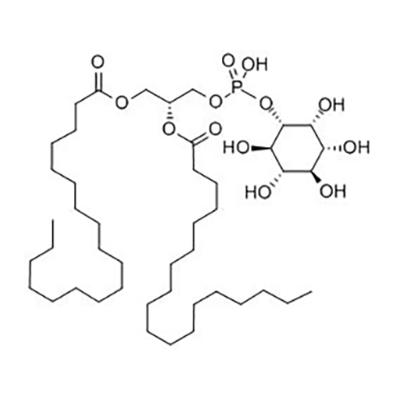
-
磷脂酰肌醇 编辑
中文名:磷脂酰肌醇
外文名:Phosphatidylinositol
化学式:C45H87O13P
CAS登录号:97281-52-2
简称:PI
组成:磷酸1,2-二脂酰甘油和肌醇
分子式:C45H87O13P
CAS号:97281-52-2
“磷脂酰肌醇(phosphatidylinositols)”是磷脂酸中的磷酸部分与肌醇酯化所产生的酰基衍生物。具体化合物的名称可以俗名磷脂酸(sn-甘油-3-磷酸)为母体进行半系统命名或按一般规则进行系统命名。
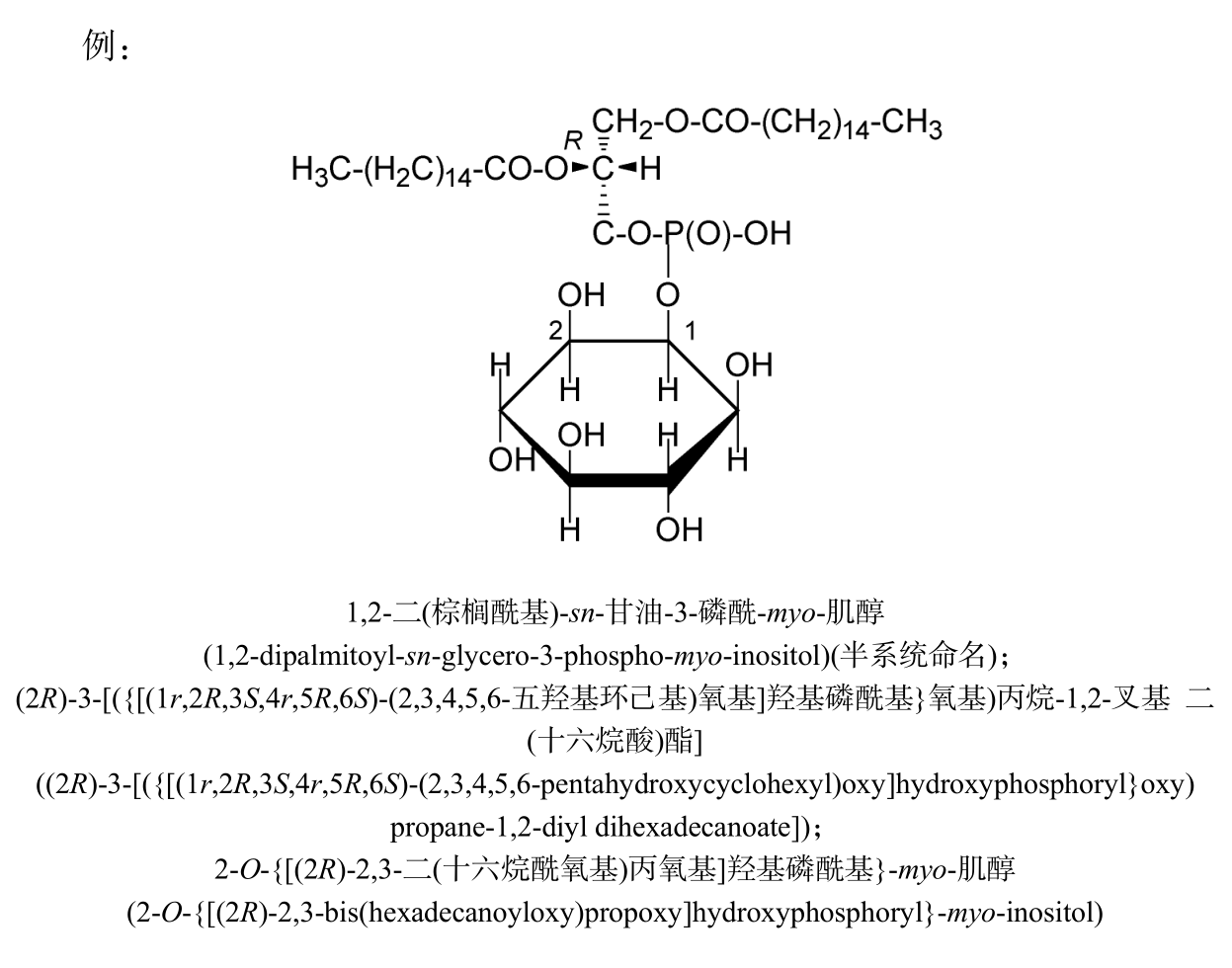 磷脂酰肌醇
磷脂酰肌醇
化学途径
是G蛋白偶联受体的信号转导通路中的一种途径, 在信号通路中胞外信号分子与细胞表面G蛋白耦联型受体结合,激活质膜上的磷脂酶C(PLC-β),使质膜上4,5-二磷酸磷脂酰肌醇(PIP2)水解成1,4,5-三磷酸肌醇(IP3)和二酰基甘油(DG)两个第二信使,胞外信号转换为胞内信号,这一信号系统又称为“双信使系统”(Double Messenger System)。
 磷脂酰肌醇
磷脂酰肌醇
DG结合于质膜上,可活化与质膜结合的蛋白激酶C(protein Kinase C,PKC)。PKC以非活性形式分布于细胞溶质中,当细胞接受刺激,产生IP3,使Ca2+浓度升高,PKC便转位到质膜内表面,被DG活化,PKC可以使蛋白质的丝氨酸/苏氨酸残基磷酸化使不同的细胞产生不同的反应,如细胞分泌、肌肉收缩、细胞增殖和分化等。DG的作用可用佛波醇酯(phorbol ester)模拟。
DG通过两种途径终止其信使作用:一是被DG-激酶磷酸化成为磷脂酸,进入磷脂酰肌醇循环;二是被DG酯酶水解成单酯酰甘油。由于DG代谢周期很短,不可能长期维持PKC活性,而细胞增殖或分化行为的变化又要求PKC长期活性所产生的效应。现发现另一种DG生成途径,即由磷脂酶催化质膜上的磷脂酰胆碱断裂产生的DG,用来维持PKC的长期效应。
首先由激活的SrcPrK和ZAP-70通过LAT使膜结合的磷脂酶C(PLC)分子丁链上的酪氨酸残基发生磷酸化。磷酸化的PLC—γ发挥酶活性,使底物二磷酸磷脂酰肌醇(PIP2)水解成两个成分:三磷酸肌醇(IP3)和二酰甘油(DAG)。IP3可迅速地从膜内侧向胞质溶胶中扩散,一方面打开细胞膜上的钙通道使Ca2+进入细胞内,同时开启细胞内钙池(内质网)增加Ca2+—的释放,协同提高胞内游离钙的浓度。胞质Ca2+含量的上升,激活一种称为钙调蛋白(camodulin)的Ca2+结合蛋白,后者可调节其他酶类的活性,并最终导致钙调磷酸酶的激活。
 T细胞抗原激活信号转导磷脂酰肌醇途径
T细胞抗原激活信号转导磷脂酰肌醇途径
钙调磷酸酶是一种丝、苏氨酸磷酸酶而不是PTK。另一方面,与胞膜内侧相联的DAG则直接激活PKC。后面熔会捍到,钙调磷酸酶和PKC主要分别活化两种重要的转录因子NF—AT和NF—cB。因而在这一条信号转导的下游通路中,实际上再一分为二,形成钙调磷酸酶参与的途径。和PKC介导的途径。由于一个PLCγ分子可以产生很多的IP2和DAG,这就放大了传入的抗原识别信号.并保证其转导的有效性。
Ca2+活化各种Ca2+结合蛋白引起细胞反应,钙调素(calmodulin,CaM)由单一肽链构成,具有四个钙离子结合部位。结合钙离子发生构象改变,可激活钙调素依赖性激酶(CaM-Kinase)。细胞对Ca2+的反应取决于细胞内钙结合蛋白和钙调素依赖性激酶的种类。如:在哺乳类脑神经元突触处钙调素依赖性激酶Ⅱ十分丰富,与记忆形成有关。该蛋白发生点突变的小鼠表现出明显的记忆无能。
IP3信号的终止是通过去磷酸化形成IP2,或被磷酸化形成IP4。Ca2+由质膜上的Ca2+泵和Na+-Ca2+交换器将抽出细胞,或由内质网膜上的钙泵抽进内质网。