-
核糖核酸 编辑
概述
 核糖核酸
核糖核酸
人体一个细胞含RNA约10pg(含DNA约7pg)。与DNA相比,RNA种类繁多,分子量较小,含量变化大。RNA可根据结构和功能的不同分为信使RNA和非编码RNA。非编码RNA分为非编码大RNA和非编码小RNA。非编码大RNA包括核糖体RNA、长链非编码RNA。非编码小RNA包括转移RNA、核酶、小分子RNA等。小分子RNA(20~300nt)包括 miRNA、 SiRNA、 piRNA、scRNA、 snRNA、 sNORNA等,细菌也有小分子RNA(50~500nt)。
名称 | 功能 | 存在 |
|---|---|---|
信使RNA(mRNA) | 所有的生物 | |
转移RNA(tRNA) | 携带氨基酸,参与翻译。 | 所有的生物 |
核糖体RNA(rRNA) | 核糖体组分,参与翻译。 | 所有的生物 |
核小RNA(snRNA) | ||
核仁小RNA(snoRNA) | 参与古菌和真核生物rRNA前体的后加工。 | 真核生物和古菌 |
微RNA(microRNA或miRNA) | 绝大多数真核生物 | |
增强子RNA(eRNA) | 真核生物 | |
小干扰RNA(siRNA) | 主要在翻译水平上抑制特定基因的表达。 | 真核生物 |
小激活RNA(saRNA) | “瞄准”特定基因的启动子,激活它们的转录。 | 某些真核生物 |
piRNA或piwi RNA | ||
长非编码RNA(lncRNA) | 在基因表达的多个环节调节基因的表达。 | 真核生物 |
7SL RNA | 作为SRP的一部分,参与蛋白质的定向和分泌。 | 真核生物和古菌 |
7SK RNA | 抑制RNA聚合酶Ⅱ催化的转录延伸。 | 脊椎动物 |
RMRP RNA | 参与线粒体DNA复制过程中RNA引物的加工;参与rRNA的后加工;参与切除一种阻滞细胞周期的蛋白质的mRNA的5'非翻译序列,而促进细胞周期的前进。 | 真核生物 |
转移信使RNA(tmRNA) | 细菌 | |
crRNA | 锁定外来核酸,引导Cas蛋白将外来核酸水解。 | 绝大多数原核生物 |
tracrRNA | 与crRNA结合,引导Cas蛋白。 | 绝大多数原核生物 |
向导RNA(gRNA) | 参与锥体虫线粒体mRNA的编辑。 | 某些真核生物 |
类病毒 | 植物 | |
端聚酶RNA | 作为端聚酶的模板,有助于端粒DNA的完整。 | 真核生物 |
核开关或RNA开关(riboswitch) | 在转录或翻译水平上调节基因的表达。 | 原核生物和少数低等的真核生物 |
核酶 | 原核或真核生物以及某些RNA病毒 | |
环状非编码RNA(circRNA) | 作为竞争性内源RNA,参与调控细胞内特定miRNA的功能;还可与细胞内一些RNA结合蛋白结合,调节这些蛋白质与其他RNA之间的相互作用。 | 主要是真核生物 |
Xist RNA | 雌性哺乳动物 |
信使RNA
信使RNA(mRNA)最早发现于1960年,在蛋白质合成过程中负责传递遗传信息、直接指导蛋白质合成,具有以下特点。
1.含量低,占细胞总RNA的1%~5%。
2.种类多,可达105种。不同基因表达不同的mRNA。
3.寿命短,不同mRNA指导合成不同的蛋白质,完成使命后即被降解。细菌mRNA的平均半衰期约为1.5分钟。脊椎动物mRNA的半衰期差异极大,平均约为3小时。
4.长度差异大哺乳动物mRNA长度为5×102~1×105nt原核生物与真核生物的mRNA虽然在结构上有差异,但功能一样,都是指导蛋白质合成的模板。
转移RNA
") 蟾蜍血涂片(用吡罗红甲基绿染色液染色)
蟾蜍血涂片(用吡罗红甲基绿染色液染色)
真核生物90%的RNA分布在细胞质中,少量存在于线粒体、叶绿体和核仁中。
原核生物的RNA分布在细胞质中。
 核糖核酸
核糖核酸
RNA和DNA一样,也是由各种核苷酸通过3′,5′-磷酸二酯键连接构成的多核苷酸链,但与DNA有一系列差异。
1.在化学组成方面,RNA含核糖而不含脱氧核糖。含尿嘧啶而不含胸腺密啶。例外的是,每个tRNA分子含有一个胸腺嘧啶,这是在RNA链合成后由尿嘧啶甲基化生的,此外,前面已提到,少数DNA含有少量核糖,但这些个别的例外并不能以此否定两类核酸组成上的差异。
2.RNA一级结构的概念虽与DNA相同.但其基本结构单位是核糖核苷酸而不是脱氧核糖核苷酸。此外,部分RNA5′端或3′端有特殊的核苷酸序列,而且RNA一级结构中没有DNA那样复杂的顺序组织。
3.绝大多数RNA为单链分子,单链可自身折迭形成发夹(hairpin)样结构而有局部双螺旋结构的特征,这是各种RAN空间结构的共同特征。RNA局部双螺旋结构中碱基互补配对规律是A对U和G对C。由于RNA分子内部不能全面形成碱基配对,故其碱基克分子比A不等于U,G不等于C,不存在DNA碱基比例的Chargaff规律。
1998年,美国两位科学家安德鲁·法尔和克雷格·梅洛在《自然》杂志上共同发表了有关发现RNA(核糖核酸)干扰机制的论文,被同行称为“近一段时间以来分子生物学最激动人心的发现之一”。
安德鲁·法尔1959年出生在美国加利福尼亚州圣克拉拉县,本科在加利福尼亚大学伯克利分校主修数学,仅用3年时间就拿到了学位。1983年获美国麻省理工学院生物学博士学位。他逐渐对涉及生命奥秘的遗传学产生了兴趣,并将其作为自己终身的学术追求。
克雷格·梅洛生于1960年,他的父亲是一名古生物学家,梅洛童年时经常跟着父亲在美国西部寻找化石。
高中时代,梅洛的兴趣逐渐转移到了基因工程方面。当时科学家克隆了人类胰岛素基因,并将其DNA(脱氧核糖核酸)置入到细菌中,这样就可以人工合成无限多的胰岛素。这一成果为全球数百万糖尿病患者带来了福音。梅洛回忆说:“科学研究能够真正地对人类健康产生影响,这个想法激起了我的兴趣。”
1998年法尔和梅洛在美国卡内基学会工作期间,他们合作发现了RNA干扰机制。
安德鲁·法尔称:“克雷格和我的工作是研究为什么一些基因会停止运行,我们试图去控制它们,我们发现了一些东西可以有效地中止它们。这些基因并不能告诉你它们能做什么,所以如果你能中止它们,你就可以开始了解它们能做什么。不过,最初发现RNA现象的是一位华人学者,非常可惜,他没有进一步弄清这是为什么。”
二人发现的是一个有关控制基因信息流程的关键机制。人们的基因组通过从细胞核里的DNA向蛋白质的合成机制发出生产蛋白质的指令,这些指令通过mRNA传送。他们发现一种可以用特定基因降解mRNA的方式,在这种RNA干扰现象中,双链RNA以一种非常明确的方式抑制了基因表达。这项技术被用于全球的实验室来确定在各种病症中哪种基因起到了重要作用。
植物、动物、人类都存在RNA干扰现象,这对于基因表达的管理、参与对病毒感染的防护、控制活跃基因具有重要意义。RNA干扰是一个生物过程,在这个过程中双链RNA以一种非常明确的方式抑制了基因表达。自1998年发现以来,RNA干扰已经作为一种强大的“基因沉默”技术而出现。RNA干扰作为研究基因运行的一种研究方法已被广泛应用于基础科学,它可能在将来产生更多更新的治疗方法。科学家认为,RNA干扰技术不仅是研究基因功能的一种强大工具,不久的未来,这种技术也许能用来直接从源头上让致病基因“沉默”,以治疗癌症甚至艾滋病,在农业上也将大有可为。
mRNA
mRNA含A、U、G、C四种核苷酸,每三个相联而成一个三联体,即密码,代表一个氨基酸的信息,故按数学中排列组合法则计算,可形成43=64个不同的密码。根据实验结果,推得64个密码与氨基酸的对应关系如下表。
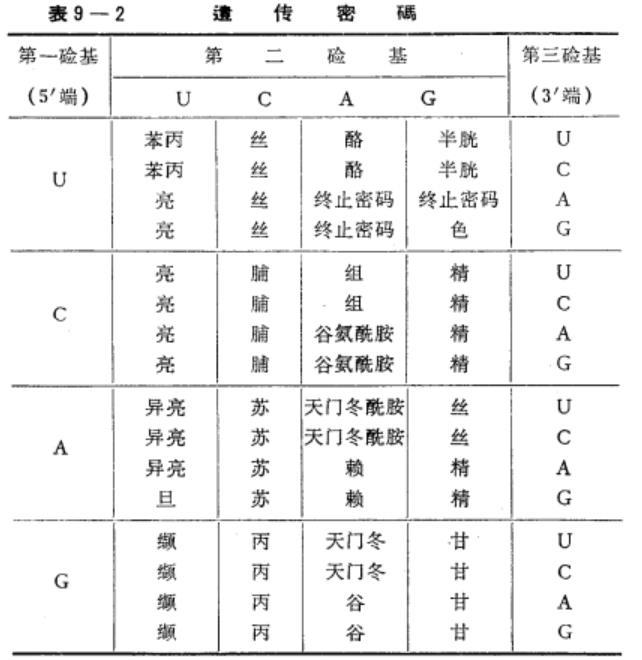 mRNA密码与氨基酸的对应关系
mRNA密码与氨基酸的对应关系
64个密码中,61个密码分别代表各种氨基酸。每种氨基酸少的只有一个密码,多的可有6个,但以2个及4个的居多数。此外,UAA、UAG、UGA这三个密码是肽链合成的终止信号,不代表任何氨基酸。在真核生物中,AUG既是甲硫氨酸的密码,又是肽链合成的起始信号;而在原核生物中,GUG(在真核生物中是缬氨酸的密码)和AUG样,都是甲酰甲硫氨酸的密码和肽链合成的起始相号。可见,除GUG外,所有的密码从细菌到高等生物都能适用,这一点为生物的共同起源学说提供了有力的佐证。
必需指出:①在mRNA整个分子中,从起始信号直至终止信号,其密码的三联体是连续的,密码与密码之间没有间隔的核苷酸;②起始信号AUG并非是mRNA的起始(5′端),而可以和5′端间隔若干个核苷酸;而且终止信号也不在mRNA的3′端。
tRNA
作为“搬运工具”的tRNA有很多种,体内20种氨基酸都有其自已特有的tRNA,所以,tRNA的种类不少于20种。tRNA在ATP供应能量和酶的作用下,可分别与特定的氨基酸结合。每个tRNA都有一个由三个核苷酸编成的“反密码”。这个反密码可以根据碱基配对的原则与mRNA上对应的密码配对,而且只有当反密码与mRNA上的密码相对应时才能配合,否则就“格格不入”。所以在翻译时,带着不同氨基酸的各个tRNA就能准确地在mRNA分子上“对号入座”,依次与典密码相合,这就保证了氨基酸能排列成一定的顺序。
tRNA上的反密码当然应能识别mRNA上相应的、互补的密码,并与之配对。然而用提纯的tRNA来进行实验时,发现一种tRNA能够识别几种密码。例如,丙氨酸tRNA,其反密码为IGC(5′>3′),可以识别三种密码。
rRNA
rRNA与多种蛋白质分子共同构成核蛋白体。核蛋白体相当于“装配机”,能促使tRNA所携带的氨基酰基缩合成肽。核蛋白体附着在mRNA上,并沿着mRNA长链的起始信号向终止信号移动。至于rRNA在蛋白质生物合成中的具体作用还不清楚。









