-
同源重组 编辑
 同源重组
同源重组
同源重组可以双向交换DNA分子,也可以单向转移DNA分子,后者又被称为基因转换(Gene Conversion)。由于同源重组严格依赖分子之间的同源性,因此,原核生物的同源重组通常发生在DNA复制过程中,而真核生物的同源重组则常见于细胞周期的S期之后。
定义
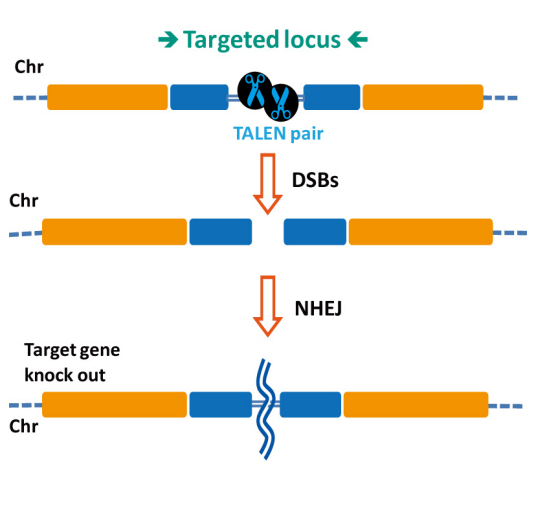
基因敲除(geneknockout),是指对一个结构已知但功能未知的基因,从分子水平上设计实验,将该基因去除,或用其它顺序相近基因取代,然后从整体观察实验动物,推测相应基因的功能。这与早期生理学研究中常用的切除部分-观察整体-推测功能的三部曲思想相似。
基因敲除可中止某一基因的表达外,还包括引入新基因及引入定点突变。既可以是用突变基因或其它基因敲除相应的正常基因,也可以用正常基因敲除相应的突变基因。
技术路线
基因敲除是80年代后半期应用DNA同源重组原理发展起来的一门新技术。80年代初,胚胎干细胞(ES细胞)分离和体外培养的成功奠定了基因敲除的技术基础。1985年,首次证实的哺乳动物细胞中同源重组的存在奠定了基因敲除的理论基础。到1987年,Thompson首次建立了完整的ES细胞基因敲除的小鼠模型。此后的几年中,基因敲除技术得到了进一步的发展和完善。
基因敲除的技术路线如下:
(1)构建重组基因载体﹔
(2)用电穿孔、显微注射等方法把重组DNA转入受体细胞核内﹔
(3)用选择培养基筛选已击中的细胞﹔
(4)将击中细胞转入胚胎使其生长成为转基因动物,对转基因动物进行形态观察及分子生物学检测。
基因敲除的靶细胞目前最常用的是小鼠ES细胞。基因敲除的技术路线虽不复杂,但由于高等真核细胞内外源DNA与靶细胞DNA序列自然发生同源重组的机率非常低,约为百万分之一,要把基因敲除成功的细胞筛选出来是一件非常困难的工作。因此,同源重组的筛选和检测就成了基因敲除技术所要解决的关键问题。目前已有多种筛选方法,其中1988年犹它大学的Capecchi教授的研究组发展的"正负双向选择系统"适用范围宽且效率较高。
同源重组(homologous recombination)是将外源基因定位导入受体细胞染色体上的方法,因为在该座位有与导入基因同源的序列,通过单一或双交换,新基因片段可替换有缺陷的基因片段,达到修正缺陷基因的目的。位点特异性重组是发生在两条DNA链特异位点上的重组,重组的发生需一段同源序列即特异性位点(又称附着点;attachmentsite,att)和位点特异性的蛋白因子即重组酶参与催化。重组酶仅能催化特异性位点间的重组,因而重组具有特异性和高度保守性。
根据氨基酸序列相似性和催化机制的差异,位点特异性重组酶主要分为两大家族,即整合酶系和解离酶/转化酶系,这两个家族相隔很远。整合酶家族催化重组的机制是进行酪氨酸介导的链交换,该家族包含有Cre/loxP、FLP/FRT等已经研究得比较清楚并广泛应用的重组酶系统。解离酶/转化酶家族催化机制是由丝氨酸介导的,当酶和DNA之间形成含磷的丝氨酸连接时,连接处DNA的4条链发生协调的交错断裂,然后重新结合,完成同源重组。该家族成员包括来自于链霉菌噬菌体的φC31整合酶、lactococcal噬菌体的TP901—1整合酶、放线菌噬菌体的R4整合酶等。中C31整合酶(φC31—int)能够有效介导噬菌体基因组中attP位点与细菌宿主染色体上attB位点间的重组反应,将外源基因整合到基因组中,使转基因的持续、高效表达,为非病毒载体转移系统在遗传病基因治疗的应用开辟了新的途径。
日本理化研究所发表新闻公报说,该所研究人员发现酵母线粒体中的DNA(脱氧核糖核酸)在一定条件下进行同源重组时,不像以前认为的那样需要DNA形成超螺旋。这一发现将为抗衰老等方面的生物医学研究提供新线索。
新闻公报说,记录生命遗传信息的DNA呈稳定的双链螺旋结构,但在复制、转录和重组等过程中,DNA链会出现一种超螺旋现象,这类似于螺旋状的电话线在受到外力时可能出现复杂的螺旋状态。
理化研究所的凌枫和柴田武彦等研究人员在酵母线粒体DNA的同源重组实验中,使用经过高度纯化的酶“Mhr1”进行催化,发现这种条件下的DNA同源重组不需要形成超螺旋,而是通过一种名为“三链体”的中间体进行。
凌枫对记者说,曾有研究证明“Mhr1”酶在抑制线粒体异质性上发挥着关键作用,此次研究揭示了其催化的反应机制核心。由于线粒体异质性与衰老等生理过程密切相关,理化研究所的这项研究为抗衰老等方面的生物医学研究提供了新线索。