-
多线染色体 编辑
各染色单体上的染色粒(见灯刷染色体)并行排列,构成多线染色体的带,带与带之间则称为间带。多线染色体的这种结构可用光学显微镜观察,也能在多线染色体上用原位分子杂交法进行基因定位,并就其结构与功能之间的关系进行系统研究,因此是细胞学和遗传学研究的有用材料。
核内DNA多次复制产生的子染色体平行排列,且体细胞内同源染色体配对,紧密结合在一起,从而阻止了染色体纤维进一步聚缩,形成体积很大的由多条染色体组成的结构叫多线染色体。多线化的细胞处于永久间期,体积也相应增大,它存在于双翅目昆虫的幼虫组织内,如唾液腺、气管等。
多线染色体不是生长到一定程度就进入有丝分裂,而是不断生长,继续复制,而且新的复制体总是沿其全长整齐地与原来的染色体并列着的,因而染色体就生长得极其庞大。例如,在果蝇唾腺细胞中每一个多线染色体都是经过大约9个循环的复制产生的,所以每条多线染色体至少包含了500-1000条单染色体(DNA纤丝),某些昆虫的多线染色体包含了多达16000条。经过醋酸洋红或地衣红染色后,在高倍光镜下就可以看到每条多线染色体都是由暗带和明间带直线交替组成的。同时也已证明,大部分DNA存在于暗区带之内,每条区带都相应于染色体上染色粒的聚合区域,它能被碱性染料染得很深,孚尔根染色呈现阳性,而明间带则几乎不着色。以后又证明了每条区带都包括几个或几十个基因位点。
1881年E.G.巴尔比安尼首先在双翅目摇蚊(ChiroNOmus)幼虫的唾腺细胞中观察到多线染色体,但未引起注意。1933年在遗传学成就的影响下T.S.佩因特在果蝇唾腺,E.海茨和H.鲍尔等在毛蚊属(Bibio)再次看到这种染色体后,人们才予重视。此后在昆虫的多种组织如肠、气管、脂肪体细胞和马尔皮基氏管上皮细胞内以及在其他动植物的一些高度特化细胞如某些原生动物及附子属(Aconitum)植物的反足细胞里也发现了这种巨大染色体。
用显微镜观察多线染色体,只须把腺体等材料放在一滴固定剂和染色剂的混合液中用玻片压破腺体细胞核,即可看到多线染色体分散出来。
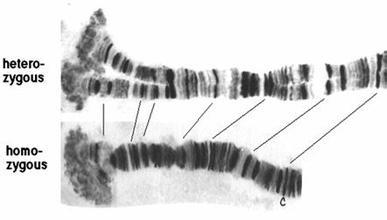
多线染色体上有按一定次序排列的横带,一个带含有多个基因。带的数目、大小、位置随不同的多线染色体而不同,因此可以据此鉴别染色体。多线染色体的带型在光镜下可见。
多线染色体上还可见膨胀而成的小泡,即疏松区。疏松区是紧密缠绕的DNA分子松开,即基因正在活动的部分,故疏松区总能检测到RNA。
并行排列的染色质纤维
 图1
图1
同种动物的不同组织以及不同动物的相同组织的多线化程度各不相同。例如摇蚊马尔皮基氏管细胞的染色体最多能复制 9次,从而使每对同源染色体组含有 512(29)条染色单体;唾腺细胞染色体复制13次,因而含有8192(213)条染色单体。
带和间带
 图2
图2
各种多线染色体上带的数目、形态、大小及其分布位置都很稳定。带和间带的分布图谱简称带谱。C.B.布里奇斯首先建立起黑腹果蝇唾腺染色体的带谱。至今已知该带的总数约有5000多条。图2示4条染色体,分别命名为X;2L(左臂)和2R(右臂);3L和3R;4。从x染色体的远端开始,依次将全部染色体分成102个区域。每一个区域可再进一步划分成A、B、C等小区域(图2)。从左边开始可将每个小区域里的带、间带和胀泡依次编号。不同带的形态和大小变化很大,同一条染色体在不同情况下其带谱不同。其他一些物种的详细带谱也有报道。
带谱与遗传图谱
把用杂交试验得到的果蝇多线染色体的遗传图谱与正常的带谱比较,可以看出每条带相当于一个遗传单位,并且可以鉴定出许多具有特殊遗传功能的带的位置。在特殊情况下,一条带可能同时有几个结构基因。例如,用原位杂交法曾经证明5SRNA基因的大部分拷贝位于2R的一条带(56F)上;组蛋白mRNA只能杂交到2L的两条带和它们之间的间带(39D-39E)上。
胀泡与基因表达
在个体发育的某个阶段或某些化学物质的诱发下,多线染色体的某些带纹变得疏松膨大而形成胀泡。最大的胀泡叫做巴尔比安尼氏环。胀泡是基因转录和翻译的形态学标志,在这里DNA解旋呈开放环,RNA的合成很活跃;核糖体排列成多聚核糖体长链,多肽链的长度有一个梯度,甚至还可观察到从巴尔比安尼环上新合成的蛋白质分泌颗粒。
在一种摇蚊 (C.pallidivittatus)唾腺前叶细胞的染色体4上,近着丝点处有巴尔比安尼环4,这些细胞能分泌特异的蛋白质颗粒。然而,在另一种摇蚊(C.tentanus)中则没有巴尔比安尼环4,也没有这种分泌颗粒。这两种动物的杂交试验表明,编码分泌颗粒蛋白质的结构基因位于染色体4的巴尔比安尼环4上。
在尖眼蕈蚊 (Sciara coprophila)等动物还发现有一种非常特殊的胀泡──DNA胀泡。3H-胸腺嘧啶核苷和3H-尿嘧啶核苷能掺入这种胀泡。它们是额外的DNA复制区域(图3),其转录活性和5-甲基胞嘧啶含量之间有反相关性。
胀泡
 多线染色体
多线染色体
激素
如果在蛋白质合成受到抑制的条件下(例如用放线菌酮等),使唾腺受到激素处理,仍能诱发早期胀泡,但不能诱发晚期胀泡。这说明早期胀泡的形成不需要蛋白质合成,晚期胀泡的形成可能是早期胀泡基因产物作用的后果。早期胀泡的活性始终依赖于蜕皮激素,一旦除去激素后胀泡就萎缩;而晚期胀泡在没有激素存在时仍能正常地出现,也说明这一情况。此外,早期和晚期胀泡对蜕皮激素浓度的反应不同,早期胀泡74EF起反应的范围很宽(可相差500倍,但晚期胀泡(62E)起反应的范围较窄(仅相差4倍)。
果蝇幼虫唾腺经高温(例如37℃,40分钟)处理后多线染色体出现明显的变化,许多胀泡萎缩,原来活跃的基因被抑制。与此同时,一组特异基因被激活,从而出现一些特异性胀泡。这一现象叫做热休克,所产生的胀泡叫做热休克胀泡(或称热诱发胀泡)。胀泡的大小与热休克处理的激烈程度有关。
果蝇多线染色体对热休克刺激的反应发生在基因表达的不同水平上。在转录(即RNA合成)水平,至少有3种反应:①诱发合成一组热休克RNA,其中有些能被翻译成热休克蛋白质;②抑制了在25℃时进行的大多数其他mRNA的合成;③中断了5S和18S+28S核糖体RNA基因转录产物的正常加工。
用3H尿苷掺入多线染色体的放射自显影试验,以及抗 RNA聚合酶Ⅱ抗体和抗染色体蛋白质的单克隆抗体在多线染色体上的分布的定位研究表明,在一般情况下,胀泡和间带是 RNA合成的活跃场所。然而经热休克处理后,RNA 聚合酶Ⅱ在转录上活跃的热休克胀泡处迅速积累起来,对染色体蛋白质专一的单克隆抗体的染色体上的分布也集中在热休克胀泡上,同时从其他区域上消失,这说明热休克胀泡是转录和转译的活跃场所。热休克处理后在转译水平上所发生的变化,一方面是诱发少数热休克多肽的合成,另一方面是抑制了原来在25℃培养时出现的大多数蛋白质的合成。热休克开始后10分钟,用35S-甲硫氨酸掺入法和SDS-聚丙烯酰胺凝胶电泳并将它们分成几条带。有关热休克多肽的确切数目,比较一致的看法是,在黑腹果蝇中主要有8种不同多肽,而在果蝇中有6种多肽。现已有直接证据表明,特异的热休克胀泡能编码特殊的热休克蛋白质。
由于多线染色体上的胀泡的变化直接反映了基因转录活性的变化,因此随着克隆重组DNA和原位分子杂交技术的发展,对多线染色体的研究必将为揭示真核生物基因的调控机制作出贡献。