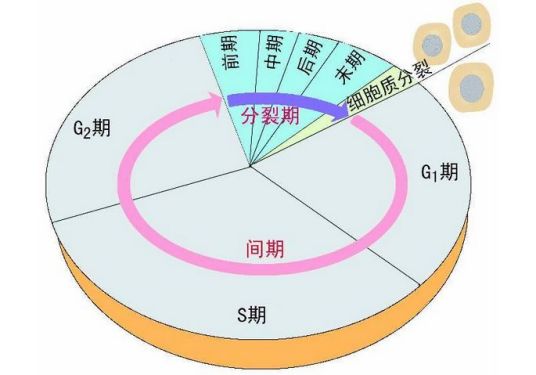
-
G2期 编辑
G2期中细胞快速生长并大量合成有丝分裂所需蛋白质。但有趣的是,G2期并不是细胞周期必需的一部分,一些细胞例如爪蟾幼胚和一些癌细胞可不经G2期而直接在DNA复制完成后进入有丝分裂。
尽管通过G2期调控了解到了基因网络的存在,但是特别是针对癌细胞还存在很多需要进一步探索的意义和控制规律。一个假说认为G2期中细胞增长是调控细胞大小的一种方法。裂殖酵母已经显示通过Cdr2介导调控Wee1活性而调整细胞大小.。虽然Wee1是相当保守的有丝分裂负向调整因子,但仍无理论阐述G2期中通用的细胞尺寸调控机制。
又名周期蛋白 B1/CDK1复合体的促成熟因子浓度达到一定程度后会使G2期结束,该复合体的活性在G2期受到严格控制。G2检查点会通过CDK1的抑制性调节来中止DNA受到损伤的细胞继续分裂。
细胞进入有丝分裂是由活性细胞周期蛋白cyclin-B1/CDK1复合物的阈值水平决定的,也被称为cyclin-B1/Cdc2或成熟促进因子(MPF)。高活性的cyclin-B1/CDK1在早期有丝分裂中引发不可逆的细胞变化,包括中心体分离、核膜破裂和纺锤体组装。在脊椎动物中,有5种cyclin B亚型(B1, B2, B3, B4和B5),但每一种亚型在调节有丝分裂进入中的具体作用尚不清楚。众所周知,细胞周期蛋白B1可以弥补细胞周期蛋白B2的损失(在果蝇中刚好相反)。酿酒酵母含有6种b型细胞周期蛋白(Clb1-6),其中Clb2是最重要的。据推测,在脊椎动物和酿酒酵母中,多种b型细胞周期蛋白的存在使得不同的细胞周期蛋白能够调控G2/M转换的不同部分,同时也使这种转换在环境扰动下更稳定。
细胞周期蛋白cyclin B1的含量在整个G1期和S期被后期促进复合物(anaphase-promoting complex APC)抑制,APC是一种E3泛素连接酶,靶向细胞周期蛋白B1,并促进其被蛋白水解。如NF-Y、FoxM1和B-Myb等转录因子被上游G1和G1/S cyclin-CDK复合物磷酸化,启动了Cyclin B1相关基因的转录,其一般开始于DNA复制后的S期末期。
在有丝分裂期间,DNA复制产生两个几乎完全相同的姐妹染色单体。DNA双链断裂发生在复制进行或G2期,可在细胞分裂发生前修复(细胞周期的m期)。因此,在G2期,一个姐妹染色单体的双链断裂可以以另一个完整的姐妹染色单体为模板进行同源重组修复
细胞在G2期对DNA损伤或不完全复制的染色体作出反应,通过延迟G2/M过渡,以防止分离受损染色体。DNA损伤可以通过激活CdC25的抑制激酶Chk1的ATM和ATR来检测。Chk1可以直接抑制Cdc25的活性,也可以通过促进Cdc2出细胞核来抑制其活性。净效应是提高细胞周期蛋白B1的阈值,从而滞后启动向m期的转变,有效地使细胞停滞在G2状态,直到通过同源定向修复等机制修复损伤。
G2到M的阻滞的长期维持也由p53介导,p53通过转录产物直接抑制Cyclin B1相关基因的表达,同时它的三个下游基因p21、GADD45、14-3-3σ也参与了G2/M期阻滞。非活化的Cyclin B1/CDK1复合物通过p21蛋白被隔离在细胞核内,而活化的Cyclin B1/CDK1复合物通过14-3-3σ蛋白被隔离在细胞质内。GADD45蛋白通过直接与cdk1相互作用抑制Cyclin B1-cdk1复合物的活性而发挥作用,14-3-3σ蛋白还可与cdc25结合,干扰Cyclin B1-cdk2复合物发挥转录调节作用。