-
免疫球蛋白A 编辑
 免疫球蛋白的5种类型
免疫球蛋白的5种类型
免疫球蛋白A单体 VS 免疫球蛋白A双体
IgA存在于两种同种型IgA1和IgA2中。它们都是高度糖基化的蛋白质。虽然IgA1在血清中占优势(~80%),但IgA2在分泌物中的含量高于血清(在分泌物中约为35%); IgA1和IgA2分泌细胞的比例在人体不同的淋巴组织中有所不同:
IgA1是血清中发现的主要IgA亚类。大多数淋巴组织具有产生IgA1的细胞的优势。
在IgA2中,重链和轻链不与二硫键连接,而是与非共价键连接。在分泌型淋巴组织(例如,肠相关淋巴组织或GALT)中,IgA2产生的份额大于非分泌性淋巴器官(例如脾,外周淋巴结)中的份额。
IgA1和IgA2都存在于外部分泌物中,如初乳,母乳,眼泪和唾液,其中IgA2比血液中更为突出。多糖抗原倾向于比蛋白质抗原诱导更多的IgA2。
IgA1和IgA2都可以是膜结合形式。
血清型免疫球蛋白A VS 分泌型免疫球蛋白A
也可以根据其位置 - 血清IgA与分泌型IgA来区分IgA的形式。
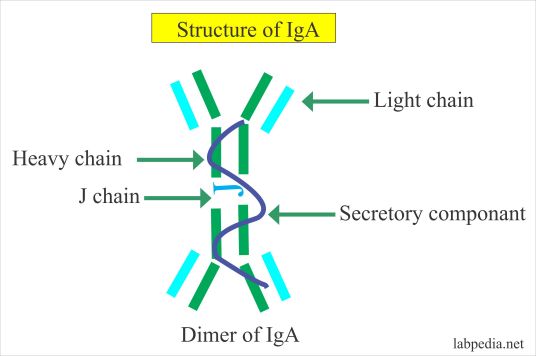
在分泌型IgA中,在分泌物中发现的形式,2-4个IgA单体的聚合物通过另外两个链连接;因此,slgA的分子量为385,000D。其中之一是J链(连接链),其是分子量为15kD的多肽,富含半胱氨酸并且在结构上与其他免疫球蛋白链完全不同。该链在IgA分泌细胞中形成。
外(粘膜)分泌物中的寡聚形式的IgA还含有分子量大得多(70kD)的多肽,称为由上皮细胞产生的分泌成分。该分子来源于聚-Ig受体(130kD),其负责寡聚(但非单体)IgA在上皮细胞上的摄取和跨细胞转运以及分泌物如泪液,唾液,汗液和肠液。
分泌型免疫球蛋白A(secretory immunoglobulin A,SIgA)是20世纪60年代初在外分泌液中发现的一种IgA抗体,主要存在于乳汁、胃肠液、呼吸道分泌液等外分泌液中。SIgA分子是由2个IgA单体(每个单体含2条轻链和2条重链)、1条J链和1条分泌片(secretory component,SC,为多聚免疫球蛋白受体的胞外裂解片段)构成的异源十聚体,为了与血清IgA单体相区别而被命名为SIgA。研究表明,SIgA是外分泌液中存在的一种主要抗体,是呼吸道、消化道、泌尿生殖道等抵御病原体及有害物质的第一道免疫防线,是机体黏膜免疫最重要的抗体。
分泌型IgA合成的相关机制
二聚体IgA(dIgA)或多聚体IgA(pIgA)从浆细胞分泌出来后,在上皮细胞的嗜碱性侧与多聚免疫球蛋白受体(polyim-munoglobulinreceptor,pIgR)以共价健形成dIgA-pIgR或pIgA-pIgR复合物,然后通过内吞作用和转运被运输到黏膜外侧,此后完整的SIgA分子通过pIgR分裂(pIgRC端跨膜部分和胞内部分在黏膜上皮细胞内降解)释放出来。SIgA在保护机体免受黏膜表面的微生物侵袭方面起着非常重要的作用,其合成与抗原提呈、淋巴细胞归巢迁移(trafficking)及周围环境中的细胞因子均有很大关系。在黏膜免疫诱导部位,抗原加工、提呈后,形成针对抗原的IgA型B细胞。在此过程中,B细胞的分化、增殖有赖T细胞的帮助。其中多种Th2样因子参与了诱导部位B细胞增殖、分化,相关因子包括TGF-β、IL-4等。前体B细胞在诱导部位内进行同种型转换(isotypeswitch),形成膜表面抗体IgA阳性的B细胞,同种型转换是形成IgA型浆细胞的关键之一。体外研究发现,在TGF-β作用下,B细胞基因重排,使Cα基因得以表达,从而使其转型为IgA型B细胞。但有研究证实IL-4的作用远高于TGF-β。在体内实验中,证实IL-4是调控B细胞在PP(潘氏结)内分化的主要因子,IL-4-/-小鼠失去合成IgA的功能,提示IL-4对于IgA的合成十分重要。
结构特征
IgA在分泌物中主要以二聚体形式存在,SIgA是由十肽组成的免疫球蛋白,来自2个不同的细胞系,沉降系数为11S,它包含2个单体的IgA、1条J链和1个分泌片,它们通过共价结合就形成所谓的SIgA。
单体IgA主要存在于血清中,含量较低,其沉降系数为7S,相对分子质量约为165×103,是重链为α的免疫球蛋白。IgA分子由2条κ链或2条λ链和2条α链构成,α链稍大于γ链。IgA经木瓜蛋白酶水解可以得到3个大小相当的片段,其中有2个相同的片段因具有抗原特异性结合能力,而被称为抗原结合片段(fragment antigen binding,Fab)。另一个片段是含有Cα2和Cα3的重链片段,它能从溶液中结晶出来,呈明显的均一性,故被称为结晶片段(fragment crystallizablc,Fc),Fc不能结合抗原,但具有各类Ig的抗原决定簇及生物活性,Ig的许多效应功能由Fc部分介导。Fab和Fc之间有一个铰链区(hingeregion),它的存在可以保证抗体分子的柔性,从而使抗体分子的许多结合位点在空间上能与抗原相互作用。人类的IgA包括IgA1和IgA2共2个亚型,它们由不同的基因表达。IgA1和IgA2的最大不同之处在于IgA1的铰链区比IgA2多13个氨基酸残基。IgA2有3种亚型,即IgA2m(1)、IgA2m(2)和IgA2n。
人类J链是相对分子质量约15×103的多肽,与其他物种的J链高度同源。人J链基因有4个外显子,外显子1编码前导肽,外显子2~4编码含有137个氨基酸残基的成熟肽,J链基因不是Ig基因簇的一部分,它定位于15号染色体。人类J链有8个半胱氨酸残基,Cys15和Cys69通过二硫键与IgA的α链相连,其他6个半胱氨酸残基形成链内二硫键(Cys13∶Cys101,Cys72∶Cys92,Cys109∶Cys134)。Johansen等发现,J链C端对于IgA聚合体的形成并非必要,但对于保持与SC的亲和力有着重要的作用;同时他们也发现,2个链内二硫键(Cys13∶Cys101和Cys109∶Cys134)对于SC的结合是不可缺少的,但对于IgA聚合体形成则可有可无,仅Cys15或Cys69的存在就足够保持多聚体IgA的稳定性。J链产生于合成IgA和IgM的浆细胞中,而且也产生于合成IgG的未成熟浆细胞,但它并不与IgG分子结合。用J链-/-鼠实验发现pIgA不能与SC结合,也不能被表达SC的上皮细胞有效转运,这说明J链参与了SC介导的转运。J链不仅是SC结合IgA的重要媒介,而且还在通过调节IgA结构而影响IgA在细胞内装配中起重要作用。
SC是上皮细胞上的pIgR的一部分,pIgR为免疫球蛋白超家族成员。pIgR由上皮细胞产生,与pIgA特别是dIgA相结合,成为IgA聚合体的转运受体,是SIgA的重要组成部分。人Fc介导子pIgA和pIgR的相互作用,pIgR细胞外部分包含5个与免疫球蛋白相似的功能域(D1~D5)。其中D1在与dIgA的Cα3功能域非共价结合过程中起了重要的作用,D5与IgA的Cα2共价结合使复合物分子更加稳定。正是SC的存在,使SIgA对蛋白酶的敏感性下降,黏液更黏稠,增强了黏附作用及防御能力。在SIgA的运输过程中,pIgR的细胞外部分与分泌性抗体结合成为固定SC,即我们经常所指的SC,可抵抗蛋白酶的降解,从而起到稳定SIgA的作用。有些未与SIgA结合的pIgR分子也被转运到黏膜外侧,并通过水解与细胞脱离,形成游离SC,与固定SC相似,亦为相对分子质量为80×103的蛋白。SC是黏膜免疫系统的重要组分,参与SIgA形成和分泌,在SC-/-转基因小鼠中,由于SC基因的缺失,不能进行pIgA的选择性上皮运输,导致该小鼠完全没有黏膜免疫功能。
功能
与普通的抗体分子相比,SIgA具有许多优良特性。SIgA分子中的J链将2个IgA单体连接起来,由于每个IgA单体具有2个抗原结合部位,因此每个SIgA抗体即有4个抗原结合位点(四价),从而比普通抗体分子具有更高的亲和力。SIgA具有很高的稳定性,其在黏膜表面的半衰期为IgG的3倍,其在人体外分泌道中的保护作用可以持续4个月以上。这种高稳定性主要是由以下几种因素所致:一是由于SIgA的铰链区较之其他抗体分子短,而铰链区是最容易受到蛋白酶攻击的部位,铰链区的缩短有利于抵抗蛋白酶的降解;二是由于SIgA的分泌片高度稳定,其多糖侧链具有防止蛋白酶降解的作用,分泌片对抗体分子的包裹使整个抗体分子变得十分稳定。此外,分泌片还赋予SIgA特殊的免疫保护作用:首先,分泌片具有非特异性的病原微生物中和活性;其次,分泌片上的糖基黏附于黏膜上皮,更使SIgA整齐地排列在黏膜表面,形成隔离保护层,可有效地阻止病毒的入侵。
血清IgA
在血液中,IgA与称为FcαRI(或CD89)的Fc受体相互作用,Fc受体在免疫效应细胞上表达,从而引发炎症反应。通过含有IgA的免疫复合物连接FcαRI引起抗体依赖性细胞介导的细胞毒性(ADCC),嗜酸性粒细胞和嗜碱性粒细胞的脱粒,单核细胞,巨噬细胞和中性粒细胞的吞噬作用,以及多形核白细胞引发的呼吸爆发活动。
分泌型IgA
粘膜区域中IgA的高患病率是产生聚合IgA(pIgA)的浆细胞和表达称为聚合Ig受体(pIgR)的免疫球蛋白受体的粘膜上皮细胞之间的合作的结果。 pIgA从附近的活化浆细胞释放并与pIgR结合。这导致IgA在粘膜上皮细胞中的转运及其从pIgR的切割以释放到外部分泌物中。
针对特定抗原的sIgA的产生取决于M细胞和潜在的树突细胞的取样,T细胞活化以及GALT,肠系膜淋巴结和小肠中的分离的淋巴滤泡中的B细胞类别转换。
聚合IgA(主要是分泌二聚体)由粘膜表面附近的固有层中的浆细胞产生。它与上皮细胞基底外侧表面的多聚免疫球蛋白受体结合,并通过内吞作用进入细胞。受体-IgA复合物在分泌到上皮细胞的腔表面上之前穿过细胞区室,仍然附着在受体上。发生受体的蛋白水解,并且二聚体IgA分子与被称为分泌成分的受体的一部分一起在整个腔内自由扩散。在肠道中,IgA可以与覆盖上皮细胞的粘液层结合。以这种方式,形成了能够在威胁到达上皮细胞之前中和威胁的屏障。
sIgA主要通过阻断上皮受体(例如通过将它们的配体结合在病原体上),通过空间阻碍与上皮细胞的附着以及通过免疫排斥来起作用。由于sIgA是一种不良的调理素和补体的激活剂,简单地结合病原体不一定足以包含它 - 特异性表位可能必须被结合以在空间上阻碍进入上皮细胞。
免疫排斥是通过将多价抗原或抗原与抗体交联,将它们捕获在粘液层中和/或清除它们的蠕动来凝集多价抗原或病原体的过程。 IgA成分的寡糖链可以与位于上皮细胞顶端的粘液层相关联。
IgA具有抗菌、抗毒、抗病毒作用,对支原体和某些真菌可能也有作用。
血清型免疫球蛋白A的作用
血清IgA具有多种抗体活性,如同种血凝素,抗胰岛素,抗布氏菌,抗白喉毒素,抗脊髓灰质炎病毒抗体等。有人认为IgA与组织抗原或蛋白抗原具有特殊结合力,从而可消除进入循环中的此类抗原,防止这些抗原诱导的炎症或自身免疫。已发现,若IgA缺乏,可伴有体内抗甲状腺球蛋白、肾上腺组织、DNA等的自身抗体水平升高。
分泌型免疫球蛋白A的作用
SIgA对机体局部免疫,如保护呼吸道、消化道粘膜有重要作用。SIgA合成功能低下的幼儿易患呼吸道或消化道感染;老年性支气管炎也可能与呼吸道SIgA合成功能降低有关。由于外分泌液中SIgA含量多,又不易被一般蛋白酶破坏,故成为抗感染、抗过敏的一道重要的免疫“屏障”。
遗传
由于遗传不能产生IgA而导致的IgA减少或缺失被称为选择性IgA缺乏,并且可以产生临床上显着的免疫缺陷。
抗IgA抗体,有时存在于IgA低或缺乏的个体中,当输入偶然含有IgA的血液制品时,可导致严重的过敏反应。然而,大多数疑似IgA过敏反应的人都经历过急性全身反应,这些反应来自抗IgA输注以外的原因。
微生物
淋病奈瑟菌(包括引起淋病的淋病奈瑟菌),肺炎链球菌和B型流感嗜血杆菌都会释放出一种破坏IgA的蛋白酶。此外,已经显示囊胚菌具有几种亚型,其产生降解人IgA的半胱氨酸和天冬氨酸蛋白酶。
自身免疫和免疫介导
IgA肾病是由肾脏中的IgA沉积物引起的。目前尚不清楚为什么IgA沉积物会发生在这种慢性疾病中。一些理论认为免疫系统的异常会导致这些沉积物。
Henoch-Schönlein紫癜(HSP)是由小血管中IgA和补体成分3(C3)沉积引起的全身性疾病。 HSP通常发生在小孩身上,涉及皮肤和结缔组织,阴囊,关节,胃肠道和肾脏。它通常伴随上呼吸道感染,并在肝脏清除IgA聚集体后几周内消退。
线性IgA大疱性皮肤病和IgA天疱疮是IgA介导的免疫性疾病的两个实例。即使使用通常有效的药物如利妥昔单抗,IgA介导的免疫性疾病通常也难以治疗。
药物性
检测方法
最常用的方法是单向免疫扩散法和免疫比浊法,但后者已逐渐代替前者。
临床意义
检测机体免疫球蛋白的含量可了解机体的体液免疫功能状态,帮助诊断免疫增生、免疫缺陷、感染及自身免疫性等多种疾病,具有重要的临床意义。
1.IgA增高:见于IgA型多发性骨髓瘤、类风湿性关节炎、系统性红斑狼疮、肝硬化及某些感染性疾病等。慢性肝病,亚急性或慢性感染性疾病(如结核、真菌感染等),自身免疫性疾病(如SLE、类风湿性关节炎),囊性纤维化,家族性嗜中性粒细胞减少症,乳腺癌,IgA肾病,IgA骨髓瘤等。
2.IgA降低:见于自身免疫病、输血反应、原发性无丙种球蛋白血症、继发性免疫缺损等。遗传性或获得性抗体缺乏症,免疫缺陷病,选择性IgA缺乏症,无γ-球蛋白血症,蛋白丢失性肠病,烧伤等。抗IgA抗体现象,免疫抑制剂治疗,妊娠后期等。
正常值参考范围
血清IgA含量正常参考值:0.7~3.8g/L。