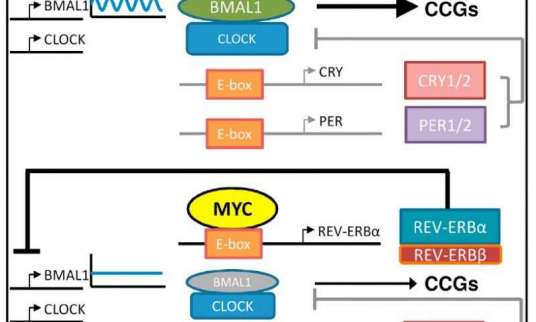
-
myc基因 编辑
myc基因首次在Burkitt淋巴瘤中发现,可通过染色体易位而活化,最常见的是通过8号染色体与14号染色体间易位,使得8号染色体上的myc基因或其相邻区域与14号染色体的免疫球蛋白重链融合而被活化。myc基因还可以通过染色体2:8或8:22间易位与免疫球蛋白轻链序列融合而被活化。尽管不同肿瘤中影响myc基因的易位断裂点的具体位置可能有所不同,但染色体易位的共通之处是改变了myc基因正常的表达调控机制。
除了染色体易位可破坏myc基因的表达调控之外,在某些肿瘤类型中myc基因还受DNA扩增的影响。myc基因在小细胞肺癌中有较高频率扩增,在很多其它类型上皮癌如乳腺癌和结直肠癌中也有扩增。
myc基因包括C - myc,N -myc,L - myc,分别定位于8号染色体,2号染色体和1号染色体。结构上由不编码蛋白质的第1外显子和编码蛋白质的第2,3外显子构成。myc基因属于编码核蛋白的癌基因,3个基因都编码一种与细胞周期调控有关的核内DNA结合蛋白。myc基因家族及其产物可促进细胞增殖,永生化,去分化和转化等,在多种肿瘤形成过程中处于重要地位。myc基因家族中的3个成员对肿瘤形成及在肿瘤类型方面存在差异。认为,C - myc的扩增与肿瘤发生与转归密切相关,N - myc的扩增对肿瘤的预后判断有意义,L - myc扩增与肿瘤的易患性和预后在不同的肿瘤中表现不一样。研究表明,myc基因产物,尤其是c - myc在诱导细胞凋亡过程中也起重要作用。
c-myc基因是禽类髓细胞病毒(AMN)MC-29的V-myc的细胞同源序列,从MC-29病毒 中分离的V-myc是gag-myc融合体,它由1358个bp的gag基因与1568个bp的V-myc基因共 同组成.C-myc基因由3个外显子及2个内含子组成,第一个外显子不编码,只起调节作 用,只有外显子2和3与V-myc相对应,编码一个439个氨基酸的蛋白质.C-myc基因由启 动子P1或P2起始转录并在第一内含子中尚有一个潜在启动子P,当第一个内含子发生 断裂时,P可被激活而成为一个异常转录起始点,但蛋白合成起始位点不变,并与正 常C-myc基因产物相同.在各种不同动物中,C-myc基因和第2.3外显子具有高度保守 性,而第1外显子则有较大的差异.小鼠和人的外显子1只有70%的同源性.人类C-myc基 因定位于8q24.在生理学上,C-myc基因的表达一般与细胞的生长状态有关,如有生长 因子刺激成纤维细胞,可导致C-myc表达增强,相反,在细胞分化时C-myc表达降低, 在细胞培养过程中,用C-myc表达结构或反义寡脱氧核酸进行研究,发现C-myc在细胞 G0期到S期的过程中也起作用.表明C-myc表达的变化与细胞的增殖及分化状态有关, 其表达产物在调节细胞生长、分化或恶性转化中发挥作用。
C-myc基因的产物为62KD的磷酸化蛋白P62c-mgc,是由C-myc基因的外显子2和3共 同编码的由439个氨基酸组成的蛋白质,定位细胞核内,为核蛋白,依C-one编码产 物,功能分类,C-myc癌基因属核蛋白基因,具有转化细胞的能力,并具有与染色体 DNA结合的特性,在调节细胞生长、分化及恶性转化中发挥作用.C-Myc蛋白在结构上 可分为转录激活区,非特异DNA结合区,核靶序列,碱性区,螺旋一环一螺旋(HLH)及 亮氨酸拉链区,在已知的转录因子中可介导蛋白的寡聚化,这两个区同时存在是 C-myc蛋白所特有的,在其它蛋白质中,很少发现.在C-Myc蛋白中,螺旋一环一螺旋 紧随着碱性区,揭示其以特异性序列方式和DNA相互作用.在以原核生物为实验对象的 研究表明,该碱性区以一个自由环存在,当以特殊方式结合到DNA上时,则变成螺 旋,该区是C-Myc蛋白与DNA特异序列的结合部位.
在C-Myc中还存在着与抑制细胞分化、自身抑制有关的区域及肿瘤转化所必需的 区域.Smith等研究了C-Myc的亮氨酸拉链区,该区介导各种转录因子的二聚作用.在亮 氨酸重复部位的突变能显蓍降低C-myc抑制鼠红白血病(MEL)细胞分化能力,同样地, 此区的插入突变能消除C-Myc的转化活性.正是这些C-myc结构成分的表达阻止了细胞 进入细胞周期,从而抑制许多细胞系的分化.Cronch等对C-Myc亮氨酸拉链区的亮氨酸 进行致突变,发现这些突变不能自身抑制,说明了亮氨酸拉链区在自身抑制中的重要 性.Stone等研究认为,C-Myc分子的中间1/3以及N-端,C-端是肿瘤转化所必需的,是 C-myc基因与肿瘤转化有关的C-myc区段.正是由于这些C-Myc功能区域的存在,从而使 C-Myc在胞浆内合成后,与其它蛋白形成寡聚体,再转移到核内,并结合到特异性的 DNA序列上,从而激活和抑制许多靶基因的转录,引起细胞生长和分化的改变,发挥 其生理调节功能及恶性转化作用.
对程序性细胞死亡Programmed CELL death. PCD)研究的深入,发现Myc蛋白参与诱导细胞凋亡.C-myc基因表达的失调是多种细胞凋亡的主要诱因,细胞发生凋亡的速度及其对诱导因素的敏感性均依赖于细胞Myc蛋白的含量.尚未成熟胸腺细胞中 Myc基因的高表达是胚胎胸腺细胞凋亡坏死的诱因.而且在细胞凋亡阶段,也观察到C-myc基因的高水平表达,如果用反义寡核苷酸阻断C-myc基因的表达,则细胞凋亡受到严重干扰.Evan研究发现,C-myc表达的失调也会启动去除生长因子后培养细胞的成熟前凋亡.他们对小鼠IL-3依赖性髓样细胞素32D进行观察,发现在洗去IL-3后, 可立即观察到C-myc基因表达下调.结果使培养细胞停止于G1期,将携带C-myc基因的 载体转染32D细胞,获得稳定表达C-myc基因的32D细胞克隆,结果这种细胞去除H-3 后,不停止于G1期,而是启动以凋亡为特征的程序性细胞死亡.结果揭示细胞凋亡是清除定点突变及细胞周期调控失衡的细胞的重要机制,一旦细胞发生障碍,C-myc基 因会启动凋零程序,相反,则导致肿瘤形成。
myc基因定位于染色体8q24、IgH、IgK、Igλ链的基因位点分别在14q32、2P13和 22q11,在BL细胞中往往出现C-myc基因位点与Ig基因位点之间的易位,即C-myc易位 到Ig位点的高活性转录区,从而组成一个高转活性的重排基因,启动C-myc转录,使 C-myc表达增强,促进细胞恶变,最后导致肿瘤的发生.
C-myc基因主要通过扩增和染色体易位重排的方式激活,与某些组织肿瘤的发 生、发展和演变转归有重要关系.在不同的人体肿瘤细胞系中,包括粒细胞性白血病 细胞系,视网膜母细胞瘤细胞系,某些神经母细胞病细胞系,乳腺癌细胞系及某些肺 癌细胞系,已发现C-myc或C-myc相关序列的扩增,在人结肠癌细胞系中也观察到 C-myc基因的扩增.C-myc癌基因已在成骨肉瘤、软骨肉瘤、脊索瘤、脂肪肉瘤、横纹 肌肉瘤中发现扩增,当扩增达到30倍时,染色体上表现HSR和DMS,而且C-myc过量表 达与肿瘤的早期复发有关,在致瘤中,已发现ras与myc、sis与myc、myc与fos偶联激 活,协同致瘤等.
许多资料表明C-myc位点在所有受检的B细胞肿瘤中有重排,Casares等发现定位 在8号染色体上的C-myc与定位在14号染色体上的Ig重链基因有同样的14.2Kb的ecori 酶切片段,因而发生8∶14易位可能是何杰氏淋巴瘤的一个普通标志.N-myc在人神经 位母细胞瘤.视网膜母细胞瘤和小细胞肺癌中有扩增,扩增的程度与肿瘤的发病进程 有关.Schwab等报告20%的神经母细胞瘤有N-myc扩增,其中大多数是侵袭性肿瘤.Seege r等报告基因拷贝数的多少预示疾病的进程,总的来说,带有一个拷贝数的Ⅰ、Ⅱ、 Ⅲ、Ⅳ.期神经母细胞瘤常规治疗效果较好,带有多个拷贝的Ⅱ、Ⅲ、Ⅳ期病人,病 情将不断加重.c-myc首先被发现与小细胞肺癌有关,30%的病例c-myc扩增,复发病人 中,myc扩增者的生存期短于没有扩增的病例,研究还发现接受化疗的肿瘤病人易引 起myc扩增.有关myc基因扩增与其它肿瘤的关系也有许多报道,认为胃癌、乳腺 癌、结肠癌、宫颈癌、何杰金氏病及头部肿瘤等都有myc基因的扩增或过度表达
过度表达,见于各种肿瘤,如肺癌,胃癌,乳腺癌,结肠癌,宫颈癌,某些神经母细胞病,粒细胞性白血病 ,视网膜母细胞瘤,成骨肉瘤,软骨肉瘤,脊索瘤,脂肪肉瘤,横纹肌肉瘤,何杰金氏病及头部肿瘤等都有myc基因的扩增或过度表达。建立同时检测Myc基因3个成员L-myc 、N-myc及C-myc异常扩增的简便方法。方法 采用聚合酶链反应(PCR)技术,用一对引物同时扩增Myc基因3个成员第2外显子中的高度保守区,扩增产物经非变性聚丙烯酰胺凝胶电泳及激光扫描。结果 此法可以检测Myc基因家族中3个成员的扩增情况。 经检测,正常组织细胞没有Myc基因扩增,32例喉癌组织中47%有L-myc和C-myc扩增,41%有N-myc 扩增,与正常比差异均有显著意义(χ=6.764,7.609, 5.961;P均<0.05)。C-myc扩增率与用PCR-琼脂糖凝胶电泳检测的结果基本相符(χ=0.254,P>0.05) 。结论 此法对检测肿瘤组织中Myc基因3个成员提供了简易、特异、 无放射污染,并且适于临床应用的、优于PCR-琼脂糖凝胶电泳的方法。
至今Myc基因已被发现了至少3种,C-myc、N-myc和L-myc
国内外用于检测N-myc、L-myc及C-myc的方法大多采用各种杂交技术。由于应用放射性同位素,需要各种防护措施,不安全又复杂,且一次只能检测Myc基因家族中的一个基因,临床上很难常规应用。PCR-琼脂糖凝胶电泳技术简单、易行、快速,但由于灵敏度低,不能分开只相差3个bpDNA的L-myc和N-myc,所以只能检测C-myc的相对扩增,而不能检测N-myc或L-myc扩增或二者的同时扩增。而且,如果有N-myc或L-myc扩增或二者同时扩增时,即使有C-myc扩增,也会出现假阴性结果。我们首先把PCR技术与中性聚丙烯酰胺凝胶电泳及激光扫描技术结合起来,并用硝酸银试剂使凝胶上的Myc 基因双链染成黑褐色。这样的实验结果特异性高,方法简单,成本低,又避免了放射性同位素的一切弊端,很适合于临床应用。聚丙烯酰胺凝胶电泳分辨率高于琼脂糖凝胶电泳,具备分离只相差一个核苷酸的不同DNA片段的特性。在中性聚丙烯酰胺凝胶中,双链DNA片段的电泳迁移率主要由DNA片段长短决定,而与其碱基组成和顺序基本无关。基于以上原理,我们设计了此实验,目的是能把琼脂糖凝胶电泳上没有分开的只相差3 bp的N-myc和L-myc分开,作到一次可同时检测3种Myc基因的扩增情况。实验表明:用PCR-中性聚丙烯酰胺凝胶电泳和用PCR-琼脂糖凝胶电泳检测C-myc基因结果基本相符, 说明本研究建立的方法结果可靠。Myc基因是较早发现的一组癌基因,Myc基因的异常扩增或表达,参与了很多肿瘤的发生和发展。本研究方法对进行3种Myc基因在肿瘤形成过程中作用机理的研究,提供了简便易行的实验手段。
1.PCR-琼脂糖凝胶电泳结果:实验设计的引物扩增Myc 3种基因,经PCR扩增,喉癌组织及正常组织细胞DNA在琼脂糖凝胶上均可见2条小于300 bp 的电泳带。第1条带为N-myc( 230bp)和L-myc(227 bp)2个DNA片段合成带,因两者只相差3 bp,故琼脂糖凝胶电泳不能将其分开而呈现1条带;第2条电泳带为C-myc基因(244 bp)。 将电泳结果拍照后的底片在激光扫描仪进行扫描,得出2条带的峰面积值。正常组织细胞的C-myc带比N-myc和L-myc合成带密度弱,比值小于1,求出正常组织细胞的C×(N+L)均值为0.6 。
C-myc扩增倍数=喉癌的C×(N+L)×0.6,考虑实验中的综合因素,定为1.5倍以上有C-myc扩增。结果显示:喉癌中有42%的C-myc扩增,正常组织细胞没有C-myc的扩增。
2.PCR-中性聚丙烯酰胺凝胶电泳结果:由于中性聚丙烯酰胺凝胶电泳对DNA的分离特性,经PCR扩增后的Myc基因电泳带显示3条。第1条带为L-myc(227 bp),第2条为N-myc(230 bp),第3条为C-myc(244 bp) 。将扩增出特异性Myc基因电泳带的凝胶片,直接在激光扫描仪上进行扫描,得出3条带的峰面积值。然后用各自的峰面积值除以各自的碱基数,作为该基因的单拷贝数,将L-myc、N-myc和C-myc三者中最小的单拷贝数设定为1,比较其他2个基因的相对倍数。正常组织细胞L-myc∶N-myc∶C-myc均值为1.0∶2.2∶3.8。结果32例喉癌中47%有L-myc和C-myc的扩增,41%有N-myc扩增。C-myc的扩增率与用PCR-琼脂糖凝胶电泳检测的结果基本一致(χ=0.254,P>0.614)。
材料
1.标本:32例喉癌组织取自我院耳鼻咽喉科住院病人,7例正常喉组织取自我科同期住院的非癌症病人,3例正常白细胞取自我院血库人全血。所有标本均经病理证实。根据喉癌组织病理分化程度不同,分为高、中、低分化癌。
2.PCR引物:5′-CCCAGCGAGACATCTGGAAGAA-3′,5′-GAGAAGCCGCTCCACATGCAGTC-3′。扩增Myc基因家族的3个基因,即C-myc(244 bp),N-myc(230 bp),L-myc(227 bp),由上海复旦大学遗传学研究所合成。
3.试剂:TaqDNA聚合酶、dNTP为Promega公司产品;蛋白酶K为Merke公司产品;硝酸银为沈阳试剂二厂产品;Marker PUC 18质粒DNA HeaⅢ酶切片段为Sigma公司产品。
方法
1.模板DNA提取:参照参考文献方法进行。
2.PCR扩增:PCR扩增总反应体积50 μl,应用模板DNA 50 ng,在DNA扩增仪操作(Perkin Elmer Cetus),循环周期为94℃1分钟,55℃1分钟,72℃2分钟,经过30个循环后,补加72℃7分钟。
3.琼脂糖凝胶电泳步骤:取PCR产物5 μl加1μl上样缓冲液(0.25%溴酚蓝,0.25%二甲苯氰FF,40%蔗糖水溶液)之后备用。制备2%琼脂糖凝胶倒入水平式电泳槽中。待胶凝固后加样,用1×TAE缓冲液作为电极缓冲液,溴化乙锭染色,电压低于5V/cm,时间约为2小时。电泳结束后,凝胶直接在紫外透射仪上观察结果。照像,拍片后的底片在激光扫描仪上进行扫描。
4.非变性聚丙烯酰胺凝胶电泳步骤:取PCR产物5 μg加1 μl上样缓冲液之后备用。制备8%非变性聚丙烯酰胺凝胶,灌注于20 cm×20 cm×0. 1 cm垂直板电泳槽中, 待凝胶聚合后加样,用1×TBE缓冲液作电极缓冲液,室温下电泳1瓦特约14小时,待指示剂溴酚蓝移到边缘时停止电泳。将电泳后的凝胶移至方盘中进行硝酸银染色。染色过程如下:用双蒸馏水(dH2O)洗凝胶3次后浸泡于染色液(硝酸银1g,加dH2O至500 ml)中30分钟,去掉染色液,用dH2O漂洗3次凝胶,然后浸泡于显色液(NaOH15g,38%甲醛3.8 ml,加dH2O至500ml)中约10分钟,当显出棕黑色DNA区带时,用dH2O洗1次凝胶后,浸于停影液(冰醋酸25 ml,加水至500 ml)中约30分钟,然后把凝胶移至固定液(乙醇25 ml,冰醋酸2.5 ml加dH2O至500 ml)中固定。固定后的凝胶在看片灯上观察结果、拍照片,凝胶直接在激光扫描仪进行扫描。
3者在第2外显子中有2个区域高度同源,3个基因分别表达各自独立的蛋白,它们的生理作用基本一致。研究表明,在许多肿瘤细胞中有Myc基因扩增或异常表达,但在不同的肿瘤组织和癌细胞系中,Myc基因家族的成员在扩增或表达方面有些差异。因此,深入研究Myc基因3个成员与人类肿瘤的关系非常有意义。本研究建立的PCR-非变性聚丙烯酰胺凝胶电泳-激光扫描技术,可同时检测肿瘤组织中3种Myc基因的扩增情况,在肿瘤发生发展过程中,为进行3种基因各自作用机理的研究提供了简易的实验手段。