-
中间纤维 编辑
 中间纤维蛋白的结构模型
中间纤维蛋白的结构模型
中间纤维具有组织特异性,不同类型细胞含有不同IF蛋白质。肿瘤细胞转移后仍保留源细胞的IF,因此可用IF抗体来鉴定肿瘤的来源。如乳腺癌和胃肠道癌,含有角蛋白,因此可断定它来源于上皮组织。大多数细胞中含有一种中间纤维,但也有少数细胞含有2种以上,如骨骼肌细胞含有结蛋白和波形蛋白。
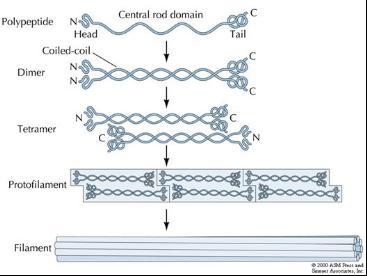
 中间纤维组装过程
中间纤维组装过程
中间纤维是细胞的第三种骨架成分,由于这种纤维的平均直径介于微管和微丝之间,故称为中间纤维。由于其直径约为10nm,故又称10nm 纤维。微管与微丝都是由球形蛋白装配起来的,而中间纤维则是由长的、杆状的蛋白装配的。中间纤维是三种骨架纤维中最复杂的一种。
与微管和微丝相比,中间纤维在结构和功能上至少有三方面的差异。首先,中间纤维是相当稳定的结构,即使用含有去垢剂和高盐溶液抽提细胞,中间纤维仍然保持完整无缺。第二,中间纤维在体积上与微管和微丝是不同的,微管是直径是24nm,微丝是7nm,而中间纤维是10nm。而且形态上也不相同,微管是由αβ微管蛋白二聚体组装成的中空管状结构,微丝是由肌动蛋白装配成的右手螺旋纤维,而中间纤维的亚基是α-螺旋杆状装配成似杆状的结构。第三,IF的亚基并不同核苷酸结合,而微管的亚基与GTP或GDP结合,微丝的亚基则与ATP或ADP结合。但是对于中间纤维装配的许多细节尚不清楚。
 中间纤维的组装模型
中间纤维的组装模型
1. 首先由平行且相互对齐的2条多肽链缠绕形成双股超螺旋二聚体(coiled-coil dimer)。此过程主要依赖于两个中间纤维蛋白单体疏水部分的结合。
2. 两个二聚体再以反向平行且端端对齐的方式组装成四聚体(tetromer),即一个二聚体的头部与另一个二聚体的尾部相连接。由于四聚体组装过程中出现了反向平行的结构特点(这与微丝和微管的组装方式不同),致使中间纤维的两端对称,从而决定了中间纤维是非极性的。
3. 每个四聚体又以头尾相连的方式延长,进一步组装成原丝(protofilament)。
4. 两根原丝平行且相互缠绕,以半分子长度交错的原则形成原纤维(protofibril),即八聚体。这种半分子长度交错排列可能是由于各种中间纤维蛋白单体头部有多精氨酸序列而中部非螺旋区L12具有多精氨酸结合位点所致。
5. 以四根原纤维互相缠绕盘曲,最终形成中间纤维。
 中间纤维的组装过程
中间纤维的组装过程
与微丝、微管不同,中间纤维蛋白合成后,基本上均装配为中间纤维,游离的单体很少。细胞内的中间纤维蛋白均受到不同程度的化学修饰,包括乙酰化、磷酸化等。
中间纤维在体外装配时不需要核苷酸和结合蛋白,也不依赖于温度和蛋白质的浓度。但在低离子强度和微碱性条件下,多数中间纤维可发生明显的解聚,一旦离子强度和pH恢复到接近生理水平时,中间纤维蛋白则迅速自我组装形成中间纤维。
在体内,大多数中间纤维蛋白都处于聚合状态,并装配形成中间纤维,很少有游离的四聚体,不存在相应的可溶性蛋白库,也没有与之平衡的踏车行为。
中间纤维结合蛋白(IF-associated protein,IFAP)是一类在结构和功能上与中间纤维密切联系,其本身又不是中间纤维结构组分的蛋白,具有很重要的功能,且有一定的细胞和组织特异性。IFAP可作为细胞中中间纤维超分子结构的调节者,中间纤维正常功能的发挥需要中间纤维结合蛋白的参与。
1. 聚纤蛋白(filaggrin) 该蛋白可结合角蛋白和波形蛋白,可使角蛋白纤维聚集形成大的纤维聚集物,因其仅在角化上皮中表达,故该蛋白的表达是角质化的分化特异性标志。
2. Triclohyallin 可束缚角蛋白使其形成紧密结构,仅在毛囊和舌上皮细胞中表达。
3. Plankin/cytolinker类中间纤维结合蛋白 其包括desmolykin、网蛋白 (pleCTin)和BPAG1(bullous PEMphigoid antigen l)三种蛋白。网蛋白参与构成桥粒和半桥粒,它还可在胞质中与中间纤维结合。BPAG1定位于内侧桥板,与角蛋白型中间纤维及其他中间纤维结合,将其固定在桥粒和半桥粒中,在桥粒和半桥粒中起着黏附和固定中间纤维的作用。
4. IFAP300 其主要功能也是与角质中间纤维结合,在桥粒和半桥粒中起着与BPAGI相同的作用,即将中间纤维锚定在桥粒上。但它在生化及免疫特性等方面与网蛋白不同。
5. 其他一些具有IFAP性质的蛋白 桥板蛋白(desmoplakin)1和2参与桥粒形成;血影蛋白及锚蛋白参与中间纤维与膜的结合;微管结合蛋白(MTP2)参与中间纤维与微管间横桥等。
由于迄今尚未找到一种对中间纤维具有特异性、可逆性影响的药物(特异性工具药),所以对中间纤维生物学功能的了解和认识并不深。随着分子生物学及分子遗传学研究方法的迅猛发展,特别是采用转基因、基因剔除等方法研究中间纤维蛋白及中间纤维结合蛋白的功能后,对中间纤维的功能有了进一步的了解。中间纤维的功能主要表现在以下几个方面。
(一)中间纤维功能的发挥具有时空特异性
中间纤维的形成及功能的发挥在不同种系的细胞及不同的发育时期均有所差异。如机体的上皮细胞可表达多种角蛋白,但在胚胎早期及成年人肝中,其上皮细胞仅表达一种Ⅰ型和Ⅱ型角质蛋白,而舌、膀胱和汗腺的上皮细胞则可表达6种甚至更多的角蛋白。在皮肤中则更加典型,不同层的上皮细胞可表达不同的角蛋白;利用这一特点,临床上可诊断肿瘤的原发部位。
(二)增强细胞的机械强度
中间纤维在受到较大的变形力时,不易断裂,中间纤维比微管和微丝更能耐受剪切力。体外实验证实上皮细胞、肌肉细胞和胶质细胞在失去完整的中间纤维网状结构后,遇到剪切力时很容易破裂。如遗传性疾病——单纯性大泡性表皮松解症患者,由于角蛋白基因突变,表达有缺陷的角质蛋白,致使表皮基底细胞中角质蛋白纤维网络被破坏,对机械性损伤非常敏感,轻微的挤压就可破坏突变的基底细胞,使患者皮肤出现水泡。表明中间纤维在提供细胞机械强度方面具有重要的意义。
(三)维持细胞和组织的完整性
中间纤维在内与核表面和核基质直接联系,在外可与细胞膜和胞外基质直接联系,其与微管、微丝及其他细胞器共同形成细胞的纤维支撑网络,可维持、固定细胞核及各种细胞器在细胞内的特定空间位置,保持细胞形态结构的完整,有利于其功能的完成。实验证实将肝细胞CK8基因剔除或转入突变的CK18基因,破坏了肝细胞中的中间纤维网状结构,结果细胞变得易破裂,最终可致肝变性、损伤、感染和坏死,表明中间纤维可维持细胞的完整性。细胞分裂后,核纤层蛋白可在内质网等一些细胞质结构的参与下形成核膜,维持细胞核的完整性。中间纤维在维持组织的完整性中也具有重要作用。
(四)与DNA复制有关
中间纤维蛋白与单链DNA之间具有高度亲和现象,提示中间纤维蛋白与DNA的复制和转录有关。核纤层蛋白在其他一些蛋白的协助下,可与染色质结合,其结合点可能是核基质黏附点、DNA复制位点和端粒。
(五)与细胞分化及细胞生存有关
在不同类型的细胞或细胞分化的不同阶段,可有不同类型的中间纤维进行特异性表达。中间纤维与组织细胞分化关系的重要例证是表皮的分化过程。表皮细胞的分化发生在最深部的生发层(基底层),伴随着细胞的分化,细胞逐渐向表皮的表层方向移动,最后形成角质细胞从表皮脱落。生发层细胞中含有前角质蛋白(prekeratin)构成的大量中间纤维束。随着细胞分化的进展,可以分别检出不同分化阶段表达出的各种角质蛋白,当细胞分化到终末阶段,细胞器及胞质中的其他蛋白均消失,只有角质蛋白中间纤维仍存在,表明它与细胞生存有关。
因为中间纤维的分布具有严格的组织特异性,所以在临床上可通过鉴定细胞中的中间纤维的类型来鉴别肿瘤细胞的组织来源及分化,确定肿瘤的性质。
(六)与细胞的信号传导有关
随着细胞内信号传导研究的深入,人们发现中间纤维在某些细胞内信号传导过程中发挥了一定的作用。