-
酿酒酵母 编辑
酿酒酵母(Saccharomyces cerevisiae),又叫面包酵母或芽殖酵母。细胞大小为2.5~10x4.5-21um。一般呈球形、卵圆形、椭圆形,有的呈圆柱状、柠檬形等。酿酒酵母细胞有两种生活形态:单倍体和二倍体。酵母单倍体的繁殖比较简单,一般是出芽生殖,当环境生存压力较大时会死亡。二倍体细胞主要进行有丝分裂繁殖,但在环境条件比较恶劣时能够以减数分裂方式繁殖,生成单倍体孢子。单倍体可以交配融合重新形成二倍体细胞,继续进行有丝分裂繁殖状态,酿酒的最适生长温度为28℃,但也可以在适当的高温下生长。作为一种单细胞真核生物,酿酒酵母具有一切真核细胞生命活动最基本特征,又有实验所需微生物应具备的背景清楚、生长迅速、操作方便等许多优点。事实上,现代遗传学、细胞生物学、生物化学中许多规律性认识都是由酵母菌为实验材料研究出来的。在这些模式酵母菌中,酿酒酵母更是世界上第一个被测出基因组DNA全序列的真核生物。它的总共6607个可读框中,已经有4752个得到了证实,这其中包括许多与细胞基本生命活动息息相关的重要基因,在结构和功能方面,也有着很强的进化保守性。至今已经发现约300种酵母蛋白质在人体中的功能相同,其中许多与人类疾病相关的蛋白相似性很高。因此,酿酒酵母一直作为一种理想生物研究模型。另外,作为传统工业发酵菌株,酿酒酵母在酒精发酵相关领域也应用广泛。因此,酿酒酵母的实用性和科研实践中的高效性都得到了充分的体现。
酵母菌的分类一直充满着挑战和争议,在分子生物学技术应用于物种分类之前,经典分类学方法主要从形态、繁殖和生理特征来进行酵母的分类,然而这些指标具有极大的局限性,酵母菌的特征可能随着培养基成分和生长阶段的改变而发生变化。截止到1998年,已描述的酵母菌达到95属,723种,目前荷兰微生物菌种保藏中心保藏有900种。常见的重要酵母菌各属有酿酒酵母属(Saccharomyces)、裂殖酵母属(Schizosaccharomyces)、汉逊酵母属(Hansenula)、毕赤酵母属(Pichia)、假丝酵母属(CandiDA)、球拟酵母属(Torulopsis)和红酵母属(Rhodotorula)等,其中酿酒酵母属是目前研究最透彻、对人类社会贡献最大的酵母属,是酿酒工业的主要菌种,还用于制造面包、糕点及医药工业等。
有关酿酒酵母属的研究可以追溯到1838年,当时Meyen首次提出了Saccharomyces这一属名,并采用双名法将酿酒酵母命名为Saccharomyces cerevisiae,Reess于1870年首次描述这一种属为具有酒精发酵能力的真菌,并将酿酒酵母命名为巴氏酵母(Saccharomyces pastorianus)。1975年,Yarrow和Nakase建立了酿酒酵母属的7种系统,之后经过多年的分子生物学鉴定和修正,直到1998年酿酒酵母属的种数被鉴定为16个,其中的酿酒酵母(S. cerevisiae)是酒精生产和果汁发酵酿酒的主要菌种。 酿酒酵母通过代谢水果、谷物、蜂蜜和其它原料中的糖产生酒精。酒精发酵以酿酒酵母(Saccharomyces cerevisiae)为主,但许多非酿酒酵母(NOn-Saccharomyces)也参与其中。 酿酒酵母也称为面包酵母或啤酒酵母,因为最早人们利用它进行啤酒和面包的工业化生产,后来人们从葡萄酒中分离培养出不同的菌株,进一步将其分为果酒用酵母、啤酒用酵母和焙烤用酵母等不同类别。
目前发现超过1500种的酵母,已鉴定700多种,但只有一少部分在工业中使用。酵母菌具有优良的发酵特性和营养特性,在实际生产中可以根据对酵母细胞数量、细胞组成成分的需求,或者对酵母代谢产物的需求,按照不同日的来确定酵母发酵的生产工艺。工业上常用的酵母种类有酿酒酵母、异常汉逊氏酵母、粟酒裂殖酵母、黏红酵母、热带假丝酵母、产朊假丝酵母、解脂假丝酵母、巴斯德毕赤酵母等。目前酵母及酵母衍生物已广泛应用于食品(酿酒、烘焙与中式发酵、调味品生产等)、医药和化工(即食营养酵母,酵母谷胱甘肽,核糖核酸、B族维生素、酵母多糖、麦角固醇等),农业(单细胞蛋白饲料、农业肥料、益生菌等)、生物能源(燃料乙醇)、生物工程(基因工程的受体菌)等领域。
1996年6月,在国际互联网的公共数据库中公布了酿酒酵母的完整基因组顺序,它被称为遗传学上的里程碑。因为首先,这是人们第一次获得真核生物基因组的完整核苷酸序列;其次,这是人们第一次获得一种易于操作的实验生物系统的完整基因组。
在酿酒酵母测序计划开始之前,人们通过传统的遗传学方法已确定了酵母中编码RNA或蛋白质的大约2600个基因。通过对酿酒酵母的完整基因组测序,发现在12068kb的全基因组序列中有5885个编码专一性蛋白质的开放阅读框。这意味着在酵母基因组中平均每隔2kb就存在一个编码蛋白质的基因,即整个基因组有72%的核苷酸顺序由开放阅读框组成。这说明酵母基因比其它高等真核生物基因排列紧密。如在线虫基因组中,平均每隔6kb存在一个编码蛋白质的基因;在人类基因组中,平均每隔30kb或更多的碱基才能发现一个编码蛋白质的基因。酵母基因组的紧密性是因为基因间隔区较短与基因中内含子稀少。酵母基因组的开放阅读框平均长度为1450bp即483个密码子,最长的是位于Ⅻ号染色体上的一个功能未知的开放阅读框(4910个密码子),还有极少数的开放阅读框长度超过1500个密码子。在酵母基因组中,也有编码短蛋白的基因,例如,编码由40个氨基酸组成的细胞质膜蛋白脂质的PMP1基因。此外,酵母基因组中还包含:约140个编码RNA的基因,排列在Ⅻ号染色体的长末端;40个编码SnRNA的基因,散布于16条染色体;属于43个家族的275个tRNA基因也广泛分布于基因组中。
序列特征
遗传信息分布在16个染色体中。其中有大约1/3的编码基因被认为是“孤儿”基因,也就是说,这些基因没有已知功能,这是因为这些基因的转录产物与酿酒酵母成其他生物所赋子功能的基因缺乏重要的同源性。这一数据仍在不断的体改中。
此染色体是由高、低G-C含量DNA结构域交替组成的,这和基因密度在染色体中的变化是呈相关性的。比如在染色体中已经证实了碱基组成的周期性变化与染色体臂重组频率的变化是平行的,富含G-C区的波峰总与每一个染色体臂中的高度重组区相重合,而富含A-T区的波谷总与低度重组的着丝点和端粒序列相重合。也有实验证实,在酵母菌中与遗传重组起始有关的基因双链分离现象,与此染色体中富含G-C的区域直接相关。
酵母基因组另一个明显的特征是含有许多DNA重复序列,其中一部分为完全相同的DNA序列,如rDNA与CUP1基因、Ty因子及其衍生的单一LTR序列等。在开放阅读框或者基因的间隔区包含大量的三核苷酸重复,引起了人们的高度重视。因为一部分人类遗传疾病是由三核苷酸重复数目的变化所引起的。还有更多的DNA序列彼此间具有较高的同源性,这些DNA序列被称为遗传冗余(genetic redundancy)。酵母多条染色体末端具有长度超过几十个kb的高度同源区,它们是遗传丰余的主要区域,这些区域至今仍然在发生着频繁的DNA重组过程。遗传冗余的另一种形式是单个基因重复,其中以分散类型最为典型,另外还有一种较为少见的类型是成簇分布的基因家族。成簇同源区(cluster homology region,简称CHR)是酵母基因组测序揭示的一些位于多条染色体的同源大片段,各片段含有相互对应的多个同源基因,它们的排列顺序与转录方向十分保守,同时还可能存在小片段的插入或缺失。这些特征表明,成簇同源区是介于染色体大片段重复与完全分化之间的中间产物,因此是研究基因组进化的良好材料,被称为基因重复的化石。染色体末端重复、单个基因重复与成簇同源区组成了酵母基因组遗传丰余的大致结构。
研究表明,遗传冗余中的一组基因往往具有相同或相似的生理功能,因而它们中单个或少数几个基因的突变并不能表现出可以辨别的表型。
 酿酒酵母的扫描电镜照片
酿酒酵母的扫描电镜照片
酿酒酵母是单细胞,卵圆形或球形,具细胞壁、细胞质膜、细胞核(极微小,常不易见到)、液泡、线粒体及各种贮藏物质,如油滴、肝糖等 。 酿酒酵母生长在麦芽汁琼脂培养基上的酿酒酵母菌落为乳白色,有光泽、平坦、边缘整齐;细胞宽度2.5-10 μm,长度 4.5 -21 μm,长与宽之比为 1 -2,多为圆形、卵圆形或卵形。
酿酒酵母细胞壁为双层结构,内层是由β-1,3-葡聚糖和 β-1,6-葡聚糖组成的葡聚糖层,在出现牙痕的附近还含有一定数量的几丁质。外层是由性质不同的各种甘露糖蛋白组成。在酵母细胞壁中,甘露糖蛋白是酵母细胞壁的重要组成部分,它除了可以参与酵母细胞之间的交配外,还与菌落形态变化以及生物大分子的免疫识别等相关。
酿酒酵母与细菌相比,在细胞大小、细胞壁组成、生长温度等方面都有很大差异(详见下表2)。
特征 | 细菌 | 酿酒酵母 |
动物肠道菌群占比 | 99% | <1% |
细胞大小/μm | 0.5-1 | 3-6 |
细胞壁组成 | 甘露糖、葡聚糖、几丁质 | |
最适生长pH | 6.5-7.5 | 4.5-6.5 |
最适生长温度/℃ | 10-80 | 20-30 |
是 | 否 | |
有 | 无 | |
肠道定植能力 | 强 | 弱 |
拮抗物质产生能力 | 强 | 弱 |
中和毒素能力 | 无 | 有 |
该表格引自:聂琴, 戴晋军, 胡骏鹏等. 酵母源生物饲料的菌种与功能. 中国饲料, 2018, (11):89-93. | ||
酿酒酵母的单倍体营养细胞和双倍体营养细胞都可以进行出芽繁殖。单倍体的营养细胞进行出芽繁殖,两个营养细胞结合,质配后进行核配,形成双倍体,进行出芽繁殖,成为双倍体的营养细胞,双倍体的营养细胞以后转变为子囊,核减数分裂形成4个子囊孢子,单倍体的子囊孢子进行芽殖。
酿酒酵母多以营养体状态进行出芽繁殖,在特定的条件下进行有性繁殖。 繁殖方式分为以下三种:
出芽繁殖:出芽时,由母细胞生出小突起,为芽体(芽孢子) 经核分裂后,一个子核移入芽体,芽体长大后与母细胞分离,单独成为新个体。繁殖旺盛时,芽体未离开母体又生新芽,常有许多芽细胞联成一串,称为假菌丝;
孢子繁殖:在不利的环境下,细胞变成子囊,内生4个孢子,子囊破裂后,散出孢子;
接合繁殖:有时每两个子囊孢子或由它产生的两个芽体双双结合成合子 合子不立即形成子囊,而产生若干代二倍体的细胞,然后在适宜区的环境下进行减数分裂,形成子囊,再产生孢子。
酿酒酵母在自然界中分布较广,属兼性厌氧微生物。繁殖时需要大量的氧气,而酒精发酵时就不需要氧气。
酿酒酵母的生长速率明显受到环境变化的影响,其中温度和pH值是主要的两个方面。温度是一种几乎可以影响细胞内所有生物化学进程的因素。高温可使一些蛋白、DNA、RNA变性,并且会影响细胞生物膜的结构和功能,而低温会抑制细胞内参加代谢反应酶的活性,从而影响酿酒酵母的生长。酿酒酵母最适生长温度为28-30℃,培养温度若高于45℃,酿酒酵母会对温度非常敏感;培养温度若在50℃持续5 min,酿酒酵母菌株99%会死亡。
大多数酿酒酵母能在pH值为2.5-8.0的培养基中生长,由于酿酒酵母有嗜酸性,根据培养温度、氧化应激、培养基和菌株的不同,最佳生长pH值约在4-6之间。培养基初始pH值过高或过低均会影响酵母菌的生长。Liu等研究发现,当pH值低于3时,酵母菌的停滞期会延长;当pH值在2.5时,酵母菌生长完全被抑制;Antonio等发现,当pH值为8时酵母菌生长速度显著降低,pH值为9时生长完全停止。
此外,氧化应激、培养基组成、渗透压等因素均会影响酿酒酵母的生长。
 干酒酿酵母
干酒酿酵母
酵母细胞原生质中含丰富的小肽,特别是谷胱甘肽(glutathione,GSH),可直接快速地被动物吸收利用,且耗能低,载体不易饱和。酿酒酵母富含核酸,其中约95%为RNA,占细胞干质量的7%-10%。
酿酒酵母中含超氧化物歧化酶(superoxide dismutase,SOD),SOD可催化超氧阴离子的歧化反应,消除超氧阴离子的毒性,对机体具有保护作用。此外,酿酒酵母菌体中还含有海藻糖、维生素、矿物质和三磷酸腺苷等营养和功能成分,这些成分也在不断地进行研究利用中。
酿酒酵母作为单细胞真核生物的代表,发酵工艺成熟,生物安全性高,主要用于燃料乙醇、白酒、葡萄酒、啤酒等的酿造生产中。 酿酒酵母活菌、非活性成分及细胞组成成分也已广泛应用于畜牧养殖业和饲料工业。
在酿酒工业上的应用
酵母菌将葡萄糖、果糖、甘露糖等单糖吸入细胞内,在无氧的条件下,经过内酶的作用,把单糖分解为二氧化碳和乙醇,此作用即发酵。
酿酒酵母乙醇生成途径:葡萄糖是很容易利用的碳源,许多微生物都能够利用葡萄糖发酵生产乙醇。酵母菌在厌氧条件下进行葡萄糖乙醇发酵,发酵过程包括葡萄糖酵解和丙酮酸的无氧降解两大生化反应过程。该过程主要由两个阶段组成,第一阶段葡萄糖通过糖酵解途径分解成丙酮酸;第二阶段丙酮酸由脱羧酶催化生成乙醛和二氧化碳,乙醛进一步被还原成乙醇。
在畜牧养殖业上的应用
 酵母菌
酵母菌
作为一种重要的模式生物,酿酒酵母可以帮助人们鉴定更多影响衰老的哺乳动物基因。由于酿酒酵母很容易进行遗传学操作和高通量筛选,它将继续作为研究人类衰老和相关疾病的理想模型。
在医药方面的应用
在医药上,因酵母富含含维生素B、蛋白质和多种酶,所以菌体制成成酵母片,用于治疗消化不良;还可从酵母菌中提取出用于生产核酸类衍生物、辅酶A.细胞色素C、谷胱甘肽和多种氨基酸的原料。
用于生产重组乙型肝炎疫苗(酿酒酵母)。该疫苗系由重组酿酒酵母表达的乙型肝炎(简称乙肝)病表面抗原( HESAR)经纯化,加入铝佐剂制成。用于预防乙型肝炎。
中美科学家针对目前已经研究清楚的四种天然苯二酚内酯聚酮化合物(monocillin II、resorcylide、lasiodiplodin和curvularin)的模式生物合成途径,利用组合生物合成技术,通过聚酮合酶亚基重排和随机组合,在酿酒酵母中异源表达新型聚酮合酶,实现一系列“非天然的”的聚酮类化合物的一步合成,为新一代药物筛选提供新的候选化合物库。
其他
酿酒酵母较其他微生物,具有良好的食品安全性;具有低pH耐受性,有利于高效积累有机酸、降低下游提取纯化成本;具有高葡萄糖耐受性,为实现高密度发酵奠定基础。酿酒酵母被认为是二羧酸生产的潜在高效细胞工厂。
单染色体酵母
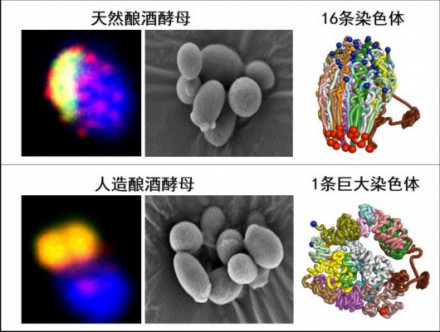
研究人员历经4年时间,通过15轮的染色体融合,最终成功创建了只有一条线型染色体的酿酒酵母菌株。经过代谢、生理、繁殖功能及染色体三维结构的鉴定,覃重军等人发现,虽然人工酵母的单条线型染色体三维结构发生了巨大变化,但这种酵母与天然酵母一样具有正常的细胞功能。
酿酒酵母菌是最简单的真核模式生物之一,具有16条染色体。中美两国的科学家都希望挑战最大限度的酵母菌染色体融合。最后,Boeke团队获得了具有2条染色体的酵母菌,而中国学者把全部16条染色体融合成了一条。
单染色体酵母的诞生,被认为是继上世纪60年代中国人工合成牛胰岛素和tRNA之后,中国在合成生物学领域的又一个重要贡献。
合成酵母
合成生物学家重建酿酒酵母的大型项目将于2020年完成。研究人员过去已成功地在许多简单的生物中完成遗传密码替代,如丝状支原体,但是要在酵母细胞中完成这项工作具有更大的挑战性,因为它们十分复杂。“合成酵母2.0”项目由四大洲的15个实验室合作展开。研究小组已经利用合成版本,替换了酿酒酵母16条染色体上的DNA。他们还试着对基因组进行重组和编辑——或删除某些片段——以了解酿酒酵母如何演化以及如何应对突变。研究人员希望经过基因工程改造的酵母细胞能够带来更高效灵活的方式,来制造更多产品,比如生物燃料和药物等。









