-
凋亡抑制基因Survivin 编辑
用免疫印迹检测方法研究Survivin基因的表达,结果发现Survivin基因在人鼻咽癌高分化上皮细胞株(CNE-1)和低分化上皮细胞株(CNE-2)中均有表达,但其表达量差异无显著意义,因此认为Survivin基因可能与鼻咽癌细胞分化程度无关,但可能与鼻咽癌的发生、发展和预后不良有关。利用间接免疫荧光、基因转染、抗体剔除(Ab-kNOck-out)、细胞平板集落形成、流式细胞术以及半胱氨酸天冬酰胺酶(caspase3)活性检测等方法,探讨EB病毒潜伏膜蛋白1(LMP1)调控细胞增殖和细胞凋亡双重效应的分子机制,发现LMP1表达介导survivin核移位,促进细胞Rb磷酸化增加,S期细胞数显著增加;LMP1通过survivin促进细胞克隆形成。用Ab-knock-out阻断survivin核移位和survivin反义核酸抑制survivin表达时,Rb磷酸化水平降低,S期细胞减少,抑制LMP1介导的细胞增殖,,活化细胞caspase3,诱导细胞凋亡。提示EB病毒LMP1可通过 添加概述促进细胞增殖和抑制细胞凋亡。
Survivin 参与细胞凋亡
细胞凋亡有两条途径:外源性凋亡途径和内源性凋亡途径。
外源性凋亡途径,又称为死亡受体通路,是由胞外肿瘤坏死因子(TNF)超家族的死亡配体如TNFa、FasL/CD95L、TWEAK和TRAIL引发的。这些配体和相关的细胞表面死亡受体(分别是TNFR、Fas/CD95、DR3、DR4/DRS)结合,使受体三聚化(receptor clustering)并激活,三聚化的死亡受体通过死亡域(deathdomain)募集衔接蛋白如TRADD和(或)FADD。衔接蛋白通过死亡效应域(deatheffeCTerdomain,DED)与 pro—caspase—8形成复合物,称为死亡诱导信号复合物(death—inducing signaling complex,DISC)。Pro-caspase-8具有弱的催化活性,在DISC中局部浓度升高,可发生自我剪接并活化。活化的caspase—8释放到胞质中启动caspase的级联反应,激活下游的效应caspase,导致细胞凋亡。激活的caspase—8能使胞质中的Bid断裂成tBid,tBid转移到线粒体上,诱导细胞色素 C从线粒体释放进入胞质,从而把死亡受体通路和线粒体通路联系起来,有效地扩大了凋亡信号。
内源性凋亡途径,又称为线粒体/细胞色素C介导的通路,线粒体是细胞生命活动控制中心,它不仅是细胞呼吸链和氧化磷酸化的中心,而且是细胞凋亡调控中心。实验表明了细胞色素C从线粒体释放是细胞凋亡的关键步骤。释放到细胞浆的细胞色素C在DATP存在的条件下能与凋亡相关因子1(Apaf-1)结合,使其形成多聚体,并促使pro-caspase-9与其结合形成凋亡小体(apoptosome),之后激活caspase-9,被激活的caspase-9能激活其它的caspase如caspase-3等,从而诱导细胞凋亡。
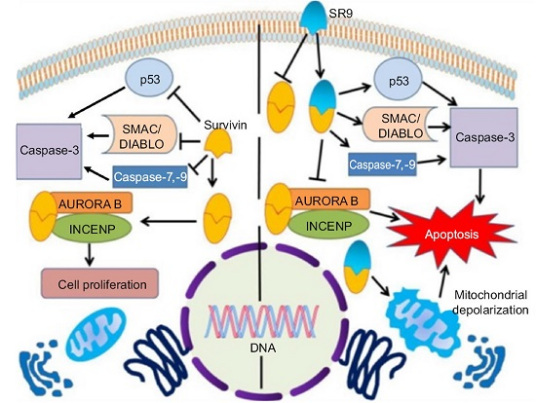
Survivin 可能主要通过两条途径来抑制细胞凋亡:①直接抑制凋亡终末效应酶Caspase-3 和Caspase-7的活性来阻断各种刺激诱导的细胞凋亡过程; ②Survivin 与周期蛋白激酶CDK4, CDK2相互作用阻断凋亡信号转导通路。
Survivin直接作用Caspase,主要通过抑制Caspasc-3和Caspasc-7的活性,Survivin也可通过P21间接抑制Caspase。Survivin与细胞周期调控因子CDK4结合, 导致 CDK2 / cyclin - E激活和核糖体(Rb)磷酸化, Rb磷酸化后启动细胞进入周期,加快G1/S期的转换,
使P21从Survivin-CDK4复合物中释放出来,与线粒体 pro-caspase-3结合,抑制caspase-3活性, 阻止线粒体释放Cyt-c,抑制细胞凋亡。一般而言,哺乳动物IAPs阻断了细胞死亡的一个普通环节,即线粒体Cyt-c的释放,直接抑制了终末效应酶Caspase-3, Caspase-7,干预了Caspase-9的活性。同时survivin抵消了一大类凋亡刺激物的作用,包括IL-3、Fas、Bax、Caspase-3、Caspase -7、Caspase -8和TNF等。