-
硝酸还原酶 编辑
真核生物中已发现三种硝酸还原酶,高等植物中最常见的一种是NADH-specific硝酸还原酶(EC1.6.6.1)。在少数几种植物中发现了第二种,即NAD(P)H-bispecific硝酸还原酶(EC1.6.6.2),它在欧洲白桦中以同源异构体式存在,在大豆和单子叶植物(玉米、水稻和大麦等)中以第二种同源异构体形式和NADH-specific硝酸还原酶一起存在。第三种形式为NADPH-specific(EC1.6.6.3),存在于真菌和苔藓中。
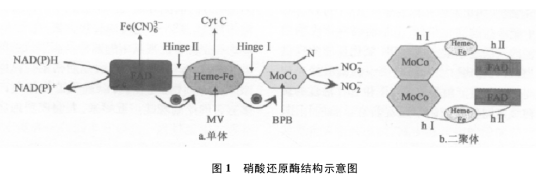
硝酸还原酶是同源二聚体,每个亚基分子质量约100~110kDA,包含三个功能区,从N-末端到C-末端分别是:钼辅酶MoCo(硝酸盐结合与降解区域),血红素-Fe(细胞色素b5结合域)和FAD(黄素腺苷酸二核苷酸磷酸、细胞色素b还原酶、NADH或NADPH结合域)。这三个区域是氧化还原中心,催化电子从NADP或NADPH转移到硝酸盐上。这三个结构和功能域通过hingeⅠ和hingeⅡ两个铰链连接起来。植物体内,hingeⅠ连接细胞色素b5结合域和钼辅酶/二聚化区域,它包含一个保守的磷酸化丝氨酸残基,被具有调控作用的14-3-3二聚体识别,在植物、真菌和哺乳动物细胞中这个二聚体来自一个高度保守的基因家族。在二价阳离子存在的情况下,hingeⅠ把硝酸还原酶转换成完全无活性的复合体,使之不能行使从NADPH到硝酸盐之间的电子转移,从而抑制酶的活性。
硝酸盐被特异性的硝酸盐转运蛋白(硝酸还原酶T)高效运输到植物根细胞中,储存在液泡里,在细胞质中被硝酸还原酶降解成亚硝酸盐。亚硝酸盐进入根的叶绿体或非绿色组织的质体中,在6-铁氧还蛋白的帮助下亚硝酸还原酶将其进一步还原成铵。细胞中发生的从硝酸盐到铵的降解过程分为两步,第一步是硝酸还原酶将硝酸盐降解成亚硝酸盐;第二步为亚硝酸还原酶将亚硝酸盐降解成铵。随后在ATP供能下,谷氨酰胺合成酶(GS)将谷氨酸和NH+4转换成谷氨酰胺,该过程是在叶绿体、细胞质或根细胞质体中进行的,最终铵参与到氨基酸的合成。谷氨酸合成酶(GOGAT)可将谷氨酰胺和α-酮戊二酸转换为两分子的谷氨酸。谷氨酸合成酶有NADH-GOGAT和Fd-GOGAT两种类型,前者主要存在于高等植物的光合细胞中,以NADH为电子供体,活性较高;后者在高等植物光合细胞和非光合细胞中均存在,以还原态的铁氧还蛋白为电子供体,活性较低。
此外,当植物细胞中NH+4浓度较高时,NADH提供电子,NH+4还可以由存在于叶绿体和线粒体中的谷氨酸脱氢酶还原为谷氨酸。但谷氨酸脱氢酶与NH+4的亲和度较低。有些氨基酸是在转氨酶的参与下由谷氨酸和酮酸参与形成的。氨基酸构成蛋白质和各种含氮化合物,也是次生化合物的起点。这里的所有酶和反应被称为硝酸盐同化途径。
硝酸还原酶、亚硝酸还原酶与N-亚硝胺的联系
植物体内的硝酸盐在硝酸还原酶和亚硝酸还原酶的作用下,通过代谢途径转化为亚硝酸盐,再转化为铵,最后被用于蛋白质的合成。土壤中硝酸盐含量过高时,植物体内的硝酸还原酶和亚硝酸还原酶不发挥作用,从而导致硝酸盐和亚硝酸盐的积累。亚硝酸盐是在叶片褐变和茎形成过程中产生的。亚硝酸盐是亚硝基化剂的前体,亚硝基化剂与烟草生物碱反应生成烟草特有亚硝胺(N-亚硝胺)。硝酸盐同化途径第二步中,亚硝酸还原酶催化亚硝酸盐还原成铵,因此,NO-3到NO-2再到NH+4的过程会因N-亚硝胺的积累而受限。通过修饰氮同化途径来控制亚硝酸还原酶活性,可影响细胞中亚硝酸盐的建成 。
植物硝酸还原酶受到很多环境因子调节,如氮源、光、O2/CO2、pH和温度,还有些内在因素如代谢物和植物激素。研究发现,将硝酸还原酶基因连接在CaMV35S启动子下构成组成型表达转入烟草中,光照或者56h黑暗处理后,转基因烟草硝酸还原酶mRNA水平高达野生型的5倍,硝酸还原酶蛋白积累,硝酸还原酶活性被激活。这些转基因植物叶片中硝酸盐的含量降低,但生长和代谢未受到明显影响。类似结果显示,硝酸还原酶或亚硝酸还原酶基因的过表达可提高mRNA水平,也提高了植物对氮的吸收,但硝酸还原酶和亚硝酸还原酶参与调控并没有促进植物生长或提高产量。通常,硝酸盐降解能力和植物生长之间没有直接联系,比如硝酸还原酶活性低的植物与野生型植株并无显著差异,但当硝酸还原酶活性比野生型低10%时,生长就受到影响。Hnsch等证明,转基因植物根部硝酸还原酶活性的丧失改变了其体内糖类和被降解的含氮物的分布,与野生型相比生长延缓。过表达硝酸还原酶的转基因烟草,吸收硝酸盐的速率比野生型慢,证明了谷氨酸或其他氨基酸的大量存在。Lea等研究表明,硝酸还原酶基因的翻译后调控可强烈影响游离氨基酸和硝酸盐的含量。一般情况下,硝酸还原酶的反常转录对代谢没有太大影响,但在翻译后水平,相关代谢产生了明显的变化。硝酸还原酶的组成型表达暂时推迟了干旱引起的硝酸还原酶活性的丧失,由此CaMV35S启动子趋动下的硝酸还原酶的表达可能促进短期缺水后氮同化获得更快的恢复 。