-
蛋白质分选 编辑
 图
图
细胞内至少存在两类蛋白质分选的信号:
①信号序列(signal sequence):存在于蛋白质一级结构上的线性序列,通常15-60个氨基酸残基,有些信号序列在完成蛋白质的定向转移后被信号肽酶(signal peptiDAse)切除.
②信号斑(signal patch):存在于完成折叠的蛋白质中,构成信号斑的信号序列之间可以不相邻,折叠在一起构成蛋白质分选的信号。
蛋白质分选信号的作用是引导蛋白质从胞质溶胶进入内质网、线粒体、叶绿体和过氧化物酶体,也可以引导蛋白质从细胞核进入细胞质或从Golgi体进入内质网。这种分选信号的氨基酸残基有时呈线性排列,有时折叠成信号斑,如引导蛋白质定向运输到溶酶体的信号斑,是溶酶体酸性水解酶被高尔基体选择性加工的标识。
一些典型的分选信号的功能及信号序列
输入细胞核
-Pro-Pro-Lys-Lys-Lys-Arg-Lys-Val-
输出细胞核
-Leu-Ala-Leu-Lys-Leu-Ala-Gly-Leu-Asp-Ile-
输入线粒体
+H3N-Met-Leu-Ser-Leu-Arg-Gln-Ser-Ile-Arg-Phe-Phe-Lys-Pro-Ala-Thr-Arg-Thr-Leu-Cys-Ser-Ser-Arg-Tyr-Leu-Leu-
输入质体
+H3N-Met-Val-Ala-Met-Ala-Met-Ala-Ser-Leu-Gln-Ser-Ser-Met-Ser-Ser-Leu-Ser-Leu-Ser-Ser-Asn-Ser-Phe-Leu- Gly-Gln-Pro-Leu-Ser-Pro-Ile-Thr-Leu-Ser-Pro-Phe-Leu- Gln-Gly-
输入过氧化物酶体
-Ser-Lys-Leu-COO-
输入内质网
+H3N-Met-Met-Ser-Phe-Val-Ser-Leu-Leu-Leu-Val-Gly-Ile-Leu-Phe-Trp-Ala-Thr-Glu-Ala-Glu-Gln-Leu-Thr-Lys-Cys- Glu-Val-Phe-Gln-
返回内质网
-Lys-Asp-Glu-Leu-COO-(KDEL)
由质膜到内体
Tyr-X-X-Φ
每一种信号序列决定特殊的蛋白质转运方向,如输入内质网的蛋白质通常N端具有一段信号序列,含有6-15个带正电荷的非极性氨基酸。由高尔基体返回内质网的蛋白质,其C端的四个氨基序列,一些已知的分选信号见表1。对于信号斑了解较少,主要是因为它存在于复杂的三维结构中,很难将其分离出来研究。
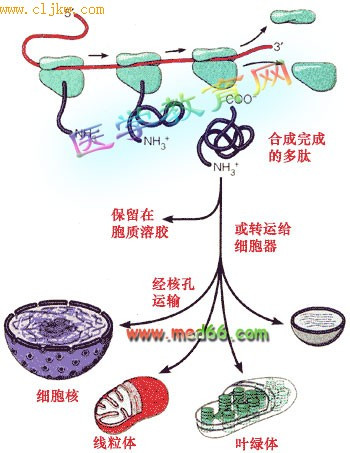
蛋白质的分选可以大体分为两条途径:
1、翻译后转运途径:在细胞质基质游离核糖体上完成多肽链的合成,然后转运至膜围绕的细胞器,如线粒体、叶绿体、过氧化物酶体及细胞核,或者成为细胞质基质的可溶性驻留蛋白和支架蛋白。
2、共翻译转运途径:蛋白质合成在游离核糖体上起始之后由信号肽引导转移至糙面内质网,然后新生肽边合成边转入糙面内质网中,再经高尔基体加工包装运至溶酶体、细胞质膜或分泌到细胞外,内质网与高尔基体本身的蛋白质分选也是通过这一途径完成的。
从蛋白质分选的转运方式和机制来看,可将蛋白质转运分为4类:
1、蛋白质的跨膜转运(transmembrane transport):主要是指在细胞质基质中合成的蛋白质转运到内质网、线粒体、质粒(包括叶绿体)和过氧化物酶体等细胞器,但进入内质网与线粒体、叶绿体和过氧化物酶体等细胞器的机制又有所不同。与分泌蛋白N端信号肽引导序列定位于内质网不同,进入线粒体、叶绿体和过氧化氢酶体等细胞器的蛋白质分选是一个多步过程,需要多个不同的寻靶序列,定位到叶绿体的前体蛋白N端具有40~50个氨基酸组成的转运肽,用以指引多肽定位到叶绿体并进一步穿透叶绿体膜进入基质中。转运到线粒体和过氧化物酶体的蛋白与此类似,但靠的是不同的引导序列,线粒体蛋白N端的导肽或过氧化物酶体蛋白C端的内在引导信号。至于这些细胞器蛋白最终是定位在不同的膜上还是不同的基质空间,除与N端不同转运肽相关外,还需要其他空间定位信号序列参与决定。此外,通过翻译后转运途径进入线粒体、叶绿体和过氧化物酶体等细胞器的蛋白质,也必须在分子伴侣的帮助下解折叠或维持非折叠状态,这有利于通过膜上的输入装置。蛋白质输入这些细胞器通常是需要能量的过程。
2、膜泡运输(vesicular transport):蛋白质通过不同类型的转运小泡从糙面内质网合成部位转运至高尔基体,进而分选转运至细胞的不同部位,其中涉及各种不同的运输小泡的定向转运,以及膜泡出芽与融合的过程。
3、选择性的门控转运(gated transport):在细胞质基质中合成的蛋白质通过核孔复合体选择性地完成核输入或丛细胞核返回细胞质(核输出),参见核孔复合体的选择性运输。
4、细胞质基质中的蛋白质转运:上述几种分选类型也涉及蛋白质在细胞基质中的转运,这一过程显然与细胞骨架系统密切相关,但由于细胞质基质的结构并不清楚,因此对其中的蛋白质转运特别是伴随信号转导途径中的蛋白质分子的转运方式了解很少。