-
生物膜 编辑
 生物膜
生物膜
其分子形态包括一个亲水性的极性头部和疏水性的脂肪酰链尾部。这种两亲性特性维持了膜结构的稳定性。亲水性头部朝向水相,疏水性尾部避水彼此聚集,这种作用称为疏水相互作用。脂质分子的双分子层排列实质上是一种熵(熵的定义式是:dS=dQ/T)的效应,满足热力学的稳定性要求,是溶液中氢键、分子间的诱导力、取向力、色散力等作用的综合结果。具有两条疏水性尾巴的磷脂分子在水相中彼此形成稳定的双分子层;对于只有一条疏水性尾巴的去垢剂、溶血磷脂等两亲性分子,则形成微团的结构;而那些尾部截面积大于头部的磷脂,则往往能形成另一种相──六角形Ⅱ相脂质的堆积特性及其形成的结构]就形成双分子层的“脂质-水”系而言,根据浓度、温度、溶液中离子种类和pH等,又会形成L(脂肪酰链呈液状自由运动的片层)、L(脂肪酰链呈直伸状且和膜面成一定倾角的片层)、L(脂肪酰链呈垂直于膜面的直伸状片层)、P(膜面呈波纹弯曲的片层)等各种相。 生物膜的脂质组成种类繁多,而且,还包含一定数量的胆固醇,所以“相”的类别多而复杂。
 生物膜
生物膜
种类 项目 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
细胞器 | 线粒体 | 叶绿体 | 高尔基体 | 内质网 | 液泡 | 溶酶体 | ||
有无膜结构 | 双层膜,分外膜和内膜 | 单层膜 | 单层膜 | 单层膜 | 单层膜 | 无 | 无 | |
主要功能 | 光合作用的场所 | |||||||
都与能量转换有关 | 都与能量转换有关 | 都与能量转换有关 | 都与能量转换有关 | 都与能量转换有关 | 都与能量转换有关 | 都与能量转换有关 | 都与能量转换有关 | 都与能量转换有关 |
完成功能的主要结构或成分 | 在内膜、基质和基粒中有许多种与有氧呼吸有关的酶 | 扁平囊和小囊泡 | 由膜构成的管道系统 | 多种水解酶 | 蛋白质和RNA | 两个相互垂直的中心粒 | ||
分布 | 所有的动植物细胞中 | 绿色植物的叶肉细胞及幼嫩茎的皮层细胞中 | 大多数动植物细胞中,一般位于核附近 | 大多数动植物细胞,广泛分布于细胞质的基质中 | 所有的植物细胞中 | 所有的动植物细胞中 | 所有的动植物细胞及原核生物中 |
 生物膜
生物膜
外周膜蛋白(peripheral membrane protein):通过与膜脂的极性头部或内在的膜蛋白的离子相互作用和形成氢键与膜的内或外表面弱结合的膜蛋白。
通道蛋白(channel protein):是带有中央水相通道的内在膜蛋白,它可以使大小适合的离子或分子从膜的任一方向穿过膜。
相关概念
 生物膜
生物膜
被动转运(passive transport):那称为易化扩散。是一种转运方式,通过该方式溶质特异的结合于一个转运蛋白上,然后被转运过膜,但转运是沿着浓度梯度下降方向进行的,所以被动转达不需要能量的支持。
主动转运(aCTive transport):一种转运方式,通过该方式溶质特异的结合于一个转运蛋白上然后被转运过膜,与被动转运运输方式相反,主动转运是逆着浓度梯度下降方向进行的,所以主动转运需要能量的驱动。在原发主动转运过程中能源可以是光,ATP或电子传递;而第二级主动转运是在离子浓度梯度下进行的。
协同运输(contransport):两种不同溶质的跨膜的耦联转运。可以通过一个转运蛋白进行同一方向(同向转运)或反方向(反向转运)转运。
胞吞(作用)(endocytosis):物质被质膜吞入并以膜衍生出的脂囊泡形成(物质在囊泡内)被带入到细胞内的过程。
细胞膜的作用和意义
细胞是人体和其他生物体一切生命活动结构与功能的基本单位。体内所有的生理功能和生化反应,都是在细胞及其合成排泄的基质(如细胞间隙中的胶原和蛋白聚糖)的物质基础上进行的。一切动物细胞都被一层薄膜所包裹,这称作细胞膜,为生物膜的一种,它把细胞内容物和细胞的周围环境分割开来。在地球上出现有生命物质和它由简单到复杂的长期演化过程中,生物膜的出现是一次飞跃,它使细胞能够既独立于环境而存在,又能通过生物膜与周围环境进行有选择的物质交换而维持生命活动。显然,细胞要维持正常的生命活动,不仅细胞的内容物不能流失,且其化学组成必须保持相对稳定,这就需要在细胞和它的环境之间有某种特殊的屏障存在。它能使新陈代谢过程中,经常由细胞得到氧气和营养物质接受各种信息分子和离子,排出代谢产物和废物,使细胞保持稳态,这对维持细胞的生命活动极为重要。因此生物膜是一个具有特殊结构和功能的选择性通透膜,它的主要功能可归纳为:能量转换、物质运送、信息识别与传递。
 生物膜
生物膜
 生物膜
生物膜
生物膜是当前分子生物学、细胞生物学中一个十分活跃的研究领域。关于生物膜的结构,生物膜与能量转换、物质运送、信息传递,以及生物膜与疾病等方面的研究及用合成化学的方法制备简单模拟膜和聚合生物膜等方面不断取得新进展。另外,人们正在研究对物质具有优良识别能力的人造膜,使模仿生物膜机能的人造内脏器官,应用于医疗诊断。
 生物膜
生物膜
物理化学特性
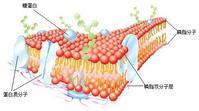
脂质的多形性 生物膜的基质是极性脂质:磷脂、胆固醇和糖脂。其分子形态包括一个亲水性的极性头部和疏水性的脂肪酰链尾部。这种两亲性特性维持了膜结构的稳定性。亲水性头部朝向水相,疏水性尾部避水彼此聚集,这种作用称为疏水相互作用。脂质分子的双分子层排列实质上是一种熵的效应,满足热力学的稳定性要求,是溶液中氢键、分子间的范德瓦耳斯力、色散力等作用的综合结果。具有两条疏水性尾巴的磷脂分子在水相中彼此形成稳定的双分子层;对于只有一条疏水性尾巴的去垢剂、溶血磷脂等两亲性分子,则形成微团的结构;而那些尾部截面积大于头部的磷脂,则往往能形成另一种相──六角形Ⅱ相(HⅡ相)(图1)。就形成双分子层的“脂质-水”系而言,根据浓度、温度、溶液中离子种类和pH等,又会形成Lα(脂肪酰链呈液状自由运动的片层)、L'(脂肪酰链呈直伸状且和膜面成一定倾角的片层)、L(脂肪酰链呈垂直于膜面的直伸状片层)、P'(膜面呈波纹弯曲的片层)等各种相。生物膜的脂质组成种类繁多,而且,还包含一定数量的胆固醇,所以“相”的类别多而复杂。
相变 脂肪酰链中的C-C单键可以旋转,产生旋转异构体。因为受到邻近基团的空间阻碍,旋转不是所有角度都能进行的。反式构象时系统的位能最小,性质最稳定;其他角度时位能都较高。一种几率较大的形式是:旋转120°后的扭转式构象。对于正丁烷,反式转为扭转式的位垒约 2.4千卡·摩尔。因而,低温时双分子层中脂肪酰链呈全反式的“僵直”状态,温度升高后链变得“柔软”。这样的转变过程不是渐行的,而是在某个温度时发生突变,该温度Tc称之为相变温度。例如DMPC(豆蔻酰磷脂酰胆碱)的Tc为23℃,DPPC(棕榈酰磷脂酰胆碱)的Tc为41℃。低于Tc时的双分子层结构称为固相或晶体相(L'、L);高于Tc时称为流动相或液晶相(Lα)。用激光拉曼光谱等方法确认了对于DPPC分子,L'-Lα相变时每条脂肪酰链大约平均新形成 6.5个扭转式键。从固相转变到流动相是个吸热的过程,相变的焓近似等于扭转式异构化所需能量与破坏相邻脂肪酰链之间的范德瓦尔斯力所需能量和脂质头部基团周围有序溶剂去结构所需能量的总和。如DPPC双分子膜,该焓值约为8.7千卡·摩尔。影响脂质分子Tc的主要因子是:①脂肪酰链的长度(长度越长,Tc越高);②脂肪酰链的饱和程度(饱和度越高,Tc越高);③脂质头部基因的种类(如,头部较小的PE(磷脂酰乙醇胺)和PC相比,Tc要高20多度。
分相
在多成分脂质系统中出现两相或更多相混合共存的状态。如在一个相当的温度区间内,固相和流动相同时存在于膜中的不同区域。分相时会影响其中膜蛋白的分布:蛋白质总是排斥于固相之外。除温度外,还有其他一些分相因子。如膜中有负电荷脂质时,介质中pH、离子种类 (特别是Ca2+)也会引起分相。L'-Lα 两相共存时,脂质双分子层的压缩率及延伸率提高,随着脂质密度涨落的出现,较大程度地提高了膜对物质的通透性。依据脂质种类和条件,也可在膜上出现双分子层和HⅡ等其他膜结构共存的分相状态。
生物膜的结构
流动镶嵌模型30年代以来,先后有许多模型用来阐述膜的结构(见细胞膜)到现在能较好地解释有关膜的各种测定数据的是1972年,S.J.辛格和G.L.尼科尔森提出的生物膜流动镶嵌模型。该模型首先根据疏水相互作用明确了双分子层中的基质是脂质,蛋白质或者靠静电相互作用结合在脂质的极性头部(外周膜蛋白),或者镶嵌在双分子层的疏水性区域(内在性膜蛋白)──此即膜的镶嵌特性。该膜型的另一要点是指出了膜的流动特性。正常生理条件下,整个脂质双分子层构成液晶状态的基质,不仅是脂质分子,蛋白质分子也处于不停的运动状态。温度、胆固醇等对膜的流动性有较大的影响。此外,脂质和蛋白质在生物膜的内、外两侧分布不对称,膜蛋白和脂质有相互作用如不少膜结合酶、抗原等都需要脂质(常是一定类型的脂质)才能表现出活性。流动镶嵌模型在某些方面还不够完善,如忽略了无机离子和水所起的作用等。
膜的流动性
脂质分子在膜中的运动形式主要有:①脂肪酰链C-C键的“反式-扭转式”异构化;②绕整个分子轴的旋转扩散;③在膜平面上的侧向扩散;④脂肪酰链的片断运动;⑤内、外层分子的翻转运动。人工膜中这种运动的几率非常小,某些生物膜中有一定几率。
膜蛋白的运动,主要是整个分子的旋转扩散及侧向扩散。此外,还存在片断运动的形式。P.G.萨夫曼和M.德尔布吕克用流体动力学方法定量表达了膜蛋白在膜上随机扩散的速率:
式中Dr为旋转扩散系数,Dl为侧向扩散系数,k为玻耳兹曼常数,T为绝对温度,μ为膜中粘滞度,μ '为外液介质的粘滞度,a为圆柱状膜蛋白的半径,h为膜的厚度,ν为欧拉常数(0.5772)。
定量测定膜流动性的方法主要有:①自旋标记法,从电子自旋共振波谱可计算出膜中标记分子的旋转相关时间(τ),但仅适用于快速运动(10-l1s<;τ<10-9s)。也可从波谱算出和脂质分子平均取向有关的参数:序参数。用饱和转移电子自旋共振波谱法则能使检测的时程扩展到10-3秒,适于对膜蛋白运动的测定。②荧光偏振法,从荧光探剂在膜中荧光的各向异性,可探测膜中的微粘滞度;而从荧光偏振的瞬态动力学则可直接测知标记分子的旋转相关时间。用闪光光解法,利用三重态荧光探剂的长寿命激发态,则能测定膜蛋白的旋转扩散。③荧光漂白恢复法,该法用以检测蛋白质、脂质分子的侧向扩散运动,适用范围是10-12cm2·s-1)<DL<10-7cm2·s-1=。
膜蛋白的限制性运动
在重建膜上,许多膜蛋白的测向扩散系数都在10-8~10-9cm2·s-1范围,和Saffman-Delbrück公式算出的理论值相符。但在生物膜上,不少膜蛋白运动很慢,甚至几乎不能运动。如红细胞膜上的带3蛋白,DL=3.8×10-l1cm2·s-1)(26℃);细菌视紫红质在嗜盐菌的紫膜上呈晶格排列,不能运动;上皮细胞类的极性细胞,其质膜的顶面区域和基底面区域上的膜蛋白种类不一样,因“紧密联结”的阻隔而不能扩散相混;LDL 受体等受体蛋白集中在特定的质膜区域──被膜穴,不能自由扩散。这些情况根据流动镶嵌模型难以解释。目前,对红细胞膜的情况有了较明确的说明:带3蛋白通过锚定蛋白 (ankyrin)和膜内侧的收缩蛋白、肌动蛋白及带4.1蛋白等组成的网络结构相联系,正是这些膜内侧的细胞骨架蛋白限制了带 3蛋白的运动(图2)。此外,尚有蛋白质彼此凝集假说、“陷阱”模型以及膜结构特殊性因素等其他解释。
生物膜的非双分子层结构 脂质双分子层是膜的基本结构,但也可能存在其他的非双分子层结构。用31P-NMR、冰冻断裂电镜术、X射线衍射等方法都表明,一些尾部截面积大于头部的脂质或带负电的脂质在一定的温度、pH、离子环境(特别是Ca2+)等条件下能形成 HⅡ相(图1)。从一些代谢活性高的内质网、线粒体、细菌质膜乃至人红细胞膜抽提出的脂质构成的膜结构中,在一定条件下都可出现HⅡ相的分子排列。活体情况下虽无HⅡ相的确切证据,但可以观察到从L相向 HⅡ相转变的过渡相──各向同性相。HⅡ相可能在膜融合、脂质分子的翻转运动及某些物质的跨膜运输等过程中起着重要的作用。
功能
物质运输 物质的跨膜运输大体可分为被动运输、主动运输和膜动运输 3大类(见生物膜离子通道)。
被动运输包括单纯扩散及促进扩散,两者都是在浓度梯度(或更广义地在电化学位梯度)的驱动下,向平衡态进行的跨膜扩散运动。用脂质分子旋转异构化所导致的“空腔”的形式和传播,可部分解释小分子、脂溶性物质的跨膜单纯扩散;而用膜中蛋白质“通道”的存在则能解释生物膜中单纯扩散的高效性,如大肠杆菌外膜中脂蛋白形成的通道、细胞之间“缝隙联结”处蛋白质形成的通道。促进扩散是膜上载体蛋白通过与被运输物质的可逆结合而促进物质的跨膜运输,表现出比单纯扩散高得多的运输速率和选择性。人红细胞膜对葡萄糖的运输、氧化磷酸化的解偶联剂对H+的运输及一些离子载体对特定离子的运输等,都属于促进扩散之列。缬氨酶素对K+的运输、尼日利亚菌素对K+/H+的交换运输都属于“移动型离子载体”。哺乳类细胞的运输系统中,膜上载体蛋白要比缬氨霉素等大得多,往往嵌入整个膜中,因此不能在膜的两侧之间来回移动。此时形成门控通道,靠蛋白质构象转换跨膜运输物质;而门控特性保证了和被运输物质的选择结合性。红细胞膜上带3蛋白对HC婣/Cl-的交换跨膜扩散即是一个很好的例子。
主动运转是物质逆着电化学位梯度跨膜运输的过程,必须有其他能量偶联输入。例如,动物细胞膜上的 Na+、K+-ATP酶靠ATP的水解,逆浓度梯度驱动Na+从细胞内向外运输,同时使K+向细胞内运输,从而维持正常生理条件下细胞内、外的 Na+、K+浓度梯度。主动运输的能量来源除 ATP外,还可来自光能、氧化磷酸化释放的能量、质子电化学位梯度以及Na+梯度等。主动运输中尚有一种在运输过程中被运输物质在膜上被转化的“基团转移”。如膜上γ-谷氨酰转肽酶使氨基酸转化成二肽,再进入细胞;细菌磷酸烯醇丙酮酸转磷酸化酶运输系统使糖转化成磷酸糖而进入细胞。
膜动运输是借膜的变形将大分子、配体、菌体等物质摄入细胞而将蛋白质、多糖等分泌出细胞的过程。其中通过膜上受体中介的内吞作用是个很重要的细胞学过程。以细胞摄入胆固醇为例:体液中的LDL(低密度脂蛋白)先和质膜上被膜穴处的LDL受体结合,然后被膜穴内凹形成被膜囊泡,在细胞内脱被膜后形成内含体,内含体很快酸性化使配体和受体解离,进而分裂成带配体及带受体的囊泡,带配体的囊泡以后和溶酶体融合。此时,LDL被水解,释放出胆固醇供细胞之用。带受体的囊泡则和质膜融合,使受体再次被利用。铁传递蛋白、胰岛素、上皮生长因子、许多毒素和病毒等亦是通过这一途径进入细胞的(见内吞与外排)。
能量转换 虽然ATP也可在可溶性酶系统中合成,但极大多数是产生在一些特定的膜上,它们称为“能量转换膜”──线粒体内膜、类囊体膜以及细菌、蓝绿藻等原核细胞的质膜。尽管这些膜在进行 ATP合成及离子运输过程中最初的能源是各种各样的,但机制却很相近。1961年P.米切尔提出“化学渗透偶联”假说,认为膜两侧H+浓度差所贮存的渗透能量能够用来产生 ATP。这一假说将膜上电子传递、离子运输及 ATP合成这三方面统一起来解释。对于线粒体,细胞呼吸时电子传递过程中游离出来的能量,以内膜两侧液相间H+的电化学位梯度(Δ)的物理能量贮存。Δ使膜上的pi+H+-ATP 酶逆转合成 ATP。植物的光合作用则是光能→渗透能→化学能。Δ包括两部分:H+的浓度差ΔpH和膜两边电位差Δψ,其关系为:
Δ=F·Δψ-2.303RTΔpH
式中F是法拉弟常数。若至少2克离子 H+的Δ合成1克分子ATP,则有关系式:
式中ΔP称为质子动力。
Δ除能用以合成 ATP外,还能作为主动运输能量、驱动细菌鞭毛的运动、产热,乃至固氮、细胞内蛋白质的运输及分泌、细胞内pH的调节等(见线粒体、叶绿体)。
信息传递 人和高等动物借助各种感受器与内外环境发生联系,这个过程和膜的可兴奋性分不开。神经细胞膜上的Na+、K+-ATP酶和Na+通道、K+通道等离子通道造成跨膜的离子浓度梯度,以及膜兴奋时Na+、K+等离子跨膜通透速率的改变,这些过程导致电兴奋延膜传递(见兴奋、神经冲动)。
细胞之间除通过物理接触直接通讯外,还能靠局部化学介体(神经生长因子、组胺等)、激素及神经递质等化学信号分子进行间接的信息传递。如肽类激素与动物细胞质膜外侧的特异性受体结合后,改变了后者的构象,在膜上作扩散运动时通过膜上G蛋白的偶联,引起膜内侧腺苷酸环化酶发生构象变化,于是催化ATP生成环腺苷酸(cAMP)。cAMP作为第二信使,激活一系列细胞内的蛋白激酶,导致众多的细胞学反应。
质膜上的钙联受体和相应的配体结合后,活化了膜上的磷脂酶C,使存在于质膜内层中的磷脂酰肌醇-4,5-二磷酸水解,形成三磷酸肌醇和甘油二酯。而后,三磷酸肌醇引起细胞内的Ca2+库(主要是内质网)释放Ca2+,于是产生一系列 Ca2+所触发的生化及细胞学反应。另一方面,甘油二酯活化质膜上的C激酶(C表示需要Ca2+来活化),使其他的一些酶磷酸化,从而产生类似cAMP的各种第二信使的效应。C激酶也能够活化膜上的 Na+/H+交换运输载体,提高细胞溶质中的pH,从而在刺激细胞生长、分化中起重要作用。
人工膜
生物膜的组分繁多,为便于研究,往往采用单一或几种脂质组成的各种人工膜结构:单分子层膜、累积膜、脂质体、平板双分子层脂膜等。另外,也可将蛋白质嵌入后组成重建膜。这些膜结构泛称“人工膜”。利用人工膜可对膜的各种物理化学特性进行深入研究。人工膜已在工程实际中得到应用,如从海水等溶液相中高效地分离和浓缩物质、利用能量转换膜原理开辟高效无污染的新能源、用作肾脏病患者的透析膜以及用于临床诊断和治疗等。
脂质体是内部为水相、由脂质双分子层形成的闭合囊泡。其种类主要有:①小单片层囊泡,大小范围为0.02~0.05微米;②多片层囊泡,大小范围为0.2~10微米;③大单片层囊泡,大小范围为0.2~10微米。除了大小、脂质成分、荷电性外,脂质体制剂尚有两个重要的参数:俘获容积和包裹效率。前者指一定量脂质体所包封的容积(摩尔/升);后者指脂质双分子层所包裹的水相所占的比例(%)。脂质体作为药物载体已用于医药临床。脂质体的水相或疏水相中包封药物后能用于癌症、溶酶体贮积病、寄生虫病、体内重金属积聚、真菌感染、关节炎等多方面疾病的治疗。这种截体系统在体内有稳定、毒性小、药物缓慢释放等优点。若膜表面加以修饰(结合抗体或特异性糖链等)或研制敏感性脂质体(在外界特定条件下膜通透性剧增的脂质体),则能使脂质体在体内具有靶向性。脂质体也可作为真核细胞的基因载体用于生物技术领域。
平板双分子层脂膜
在分隔两个水相的隔板中间若有1小孔(面积一般小于1平方厘米,则小孔处的脂滴会逐渐形成厚度只有双分子层厚的膜,此即平板双分子层脂膜(BLM)。在BLM形成过程中,脂滴厚度逐渐变薄,此时从显微镜中看到膜的颜色由各种彩色变到黑色,故BLM又称黑膜。这种人工膜最适于膜电特性的测量研究。膜中嵌入离子通道等膜蛋白后,可方便地根据测量到的电特性研究通道特性、离子通透特性、膜融合特性等。若BLM 中嵌入植物、动物以及细菌的对光敏感的色素活性物质,则可作为色素膜进行模拟研究,因为电化学测定法的灵敏度相当高,所以也可根据膜的电特性和通透特性的变化来检测环境中毒物的存在及其对机体作用的原初机制。
生物膜的化学成分
生物膜的化学成分主要有脂类、蛋白质和糖类,此外还含水、无机盐和少量的金属离子。膜中脂类和蛋白质构成了膜的主体,糖类多以复合糖的形式存在,与膜脂或膜蛋白结合分别形成膜糖脂或膜糖蛋白。
1.膜脂
构成膜的脂类有磷脂、胆固醇和糖脂,其中以磷脂为最多。这三种脂类都是双亲媒性分子,即它们都是由一个亲水的极性头部和一个疏水的非极性尾部组成。由于膜脂的这一结构特点,它们在水溶液中能自动聚拢形成脂双分子层,其游离端往往有自动闭合的趋势,形成一种自我封闭而稳定的中空结构,称脂质体。
磷脂 真核细胞膜中的磷脂主要有卵磷脂(磷脂酰胆碱)、脑磷脂(磷脂酰乙醇胺)、磷脂酰丝氨酸、鞘磷脂合磷脂酰肌醇。
胆固醇 是细胞膜内的中性脂类。真核细胞膜中胆固醇含量较高,有的膜内胆固醇与磷脂之比可达1∶1。胆固醇也是双亲媒性分子,包括三部分:极性的羟基团头部、非极性的固醇环和非极性的脂肪酸链尾部。在膜中,胆固醇分子散布在磷脂分子之间,其极性的羟基头部紧靠磷脂的极性头部,将固醇环固定在近磷脂头部的碳氢链上,其余部分分离。这种排列方式对膜的稳定性十分重要。
糖脂 是含一个或几个糖基的脂类,也是双亲媒性分子,存在于所有的动物细胞膜中,约占膜外层脂类分子的50%。动物细胞膜中的糖脂主要是鞘氨醇的衍生物,结构与鞘磷脂相似,只是其头部以糖基替代了磷脂酰碱基。脑苷脂是最简单的糖脂,只含一个糖基(半乳糖或葡萄糖)。在所有细胞中,糖脂均位于膜的非胞质面单层,并将糖基暴露在细胞表面,其作用可能是作为某些大分子的受体,与细胞识别及信息传导有关。
2.膜蛋白
生物膜所含的蛋白叫膜蛋白,是生物膜功能的主要承担者。根据蛋白分离的难易及在膜中分布的位置,膜蛋白基本可分为两大类:外在膜蛋白和内在膜蛋白。外在膜蛋白约占膜蛋白的20%~30%,分布在膜的内外表面,主要在内表面,为水溶性蛋白,它通过离子键、;氢键与膜脂分子的极性头部相结合,或通过与内在蛋白的相互作用,间接与膜结合;内在蛋白约占膜蛋白的70%~80%,是双亲媒性分子,可不同程度的嵌入脂双层分子中。有的贯穿整个脂双层,两端暴露于膜的内外表面,这种类型的膜蛋白又称跨膜蛋白。内在膜蛋白露出膜外的部分含较多的极性氨基酸,属亲水性,与磷脂分子的亲水头部邻近;嵌入脂双层内部的膜蛋白由一些非极性的氨基酸组成,与脂质分子的疏水尾部相互结合,因此与膜结合非常紧密。
 生物膜
生物膜
1.膜的流动性
 生物膜流动镶嵌模型
生物膜流动镶嵌模型
2.膜的不对称性
以脂双层分子的疏水端为界,生物膜可分为近胞质面和非胞质面内外两层,生物膜内外二层的结构和功能有很大差异,这种差异称为生物膜的不对称性。
膜脂分布的不对称主要体现在膜内外两层脂质成分明显不同。如磷脂中的磷脂酰胆碱和鞘磷脂多分布在膜的外层,而磷脂酰乙醇胺、磷脂酰丝氨酸和磷脂酰肌醇多分布在膜的内层,其中磷脂酰乙醇胺和磷脂酰丝氨酸的头部基团均带负电,致使生物膜内侧的负电荷大于外侧。膜蛋白分布的不对称主要体现在三个方面:① 即使是膜内在蛋白都贯穿膜全层,但其亲水端的长度和氨基酸的种类与顺序也不同;②外在蛋白分布在膜的内外表面的定位也是不对称的,如具有酶活性的膜蛋白Mg2+-ATP酶、5'核苷酸酶、磷酸二酯酶等均分布在膜的外表面,而腺苷酸环化酶分布在膜的内表面;③含低聚糖的糖蛋白,其糖基部分布在非胞质面。
生物膜的主要化学成分是脂类和蛋白质,还有少量糖类。关于这些组分在膜中是如何排列和组织的、以及它们之间是如何相互作用的等问题,许多学者进行了多方面的研究,先后提出了数十种不同的生物膜分子结构模型,下面介绍公认的流动镶嵌模型。
这一模型是Singer和Nicolson在1972年提出的。流动镶嵌模型保留了夹层学说和单位膜模型中磷脂双层的排列方式,即流动的脂双层分子构成膜的连续主体,蛋白质分子以不同程度镶嵌于脂质双层中。它的主要特点是:①强调了膜的流动性,膜中脂类分子既有固体分子排列的有序性,又有液体的流动性,即流动的脂类分子层构成膜的连续整体;②强调了膜的不对称性和脂类与蛋白质分子的镶嵌关系。膜中球形蛋白质分子不同程度地镶嵌在脂类双分子层中,蛋白质分子的非极性部分嵌入脂类双分子层的疏水尾部去,极性部分露于膜的表面,似一群岛屿一样,无规则地分散在脂类的海洋中。这二模型的不足之处在于它忽视了蛋白质分子对脂类分子流动性的控制作用,忽视了膜的各个部分流动性的不均匀性等等。
下面介绍几种其它模型:DAvson 和Danielli提出的 蛋白质--脂质--蛋白质 的三明治模型。
1959年,J.D.RobERTson 发展了三明治模型,提出了单位膜模型。
Simon 于1988年提出脂筏模型。
小分子物质的跨膜运输
每一个活细胞要维持其正常的生命活动,必须通过细胞膜从外界及时地吸取营养物质,同时要不断地排出其代谢产物。这些营养物质和代谢产物进出生物膜的方式,根据是否需要膜蛋白的介导分为单纯扩散和膜蛋白介导的跨膜运输两种。根据运输过程中是甭消耗代谢能又把后者分为被动运输和主动运输两种方式。
1.膜的选择性通透和单纯扩散
一些物质不需要膜蛋白的帮助,能顺浓度梯度自由扩散,通过膜的脂双层,这种跨膜运输的形式,称为单纯扩散,又称为被动扩散,它不需要消耗能量,是物质跨膜运输中最简单的一种形式。一般来说分子量小、脂溶性强的非极性的分子能迅速地通过脂双层膜,不带电荷的小分子也较易通透,如CO2、O2、乙醇和尿素可迅速扩散通过脂双层。H2O因为分子小,不带电荷,且本身具有双极结构,也很容易通过膜。一些带电分子如Na+、K+、Cl-等尽管分子很小,往往因其周围形成的水化层而难以通过脂双层的疏水区而完全不能通透。不带电的葡萄糖,因分子太大,也几乎不能自由扩散过膜。
2.膜蛋白介导的跨膜运输
对一些相对较大的极性或带电的分子,如葡萄糖、氨基酸及离子等物质均不能自由通过膜。这些物质的运输均需要有膜蛋白的介导,这些蛋白称膜运输蛋白。根据膜蛋白介导物质运输的形式,又可分为载体蛋白介导和通道蛋白介导两大类型。
跨膜运输
真核细胞中,一些大分子如蛋白质、多糖、多肽之类的物质的跨膜运输是通过细胞质膜的变形运动来完成的。这就是细胞内吞作用和细胞外吐作用。
1.内吞作用(dog)
内吞作用又称入胞作用,是通过质膜的变形运动将细胞外物质转运入细胞内的过程。根据入胞物质的不同大小,以及入胞机制的不同可将内吞作用分为三种类型:吞噬作用、吞饮作用、受体介导的内吞作用。1、吞噬作用(phagaocytosis)是指摄入直径大于1μm的颗粒物质的过程。在摄入颗粒物质时,细胞部分变形,使质膜凹陷或形成伪足将颗粒包裹摄人细胞。伪足的伸出是由肌动蛋白参与的,若用抑制肌动蛋白聚合的药物如细胞松弛素能抑制细胞吞噬。
2.吞饮作用(pinocytosis)是细胞摄入溶质或液体的过程。细胞吞饮时局部质膜下陷形成一小窝,包围液体物质,然后小窝离开质膜形成小泡,进入细胞。吞饮作用分为液相内吞和吸附内吞。前一种方式为非特异性细胞把细胞外液及其内可溶性物质摄入细胞内。后一种方式中,细胞外大分子或颗粒物质先以某种方式吸附在细胞表面,随后被摄入细胞内。如阳离子铁蛋白以静电作用先吸附在带负电荷的细胞表面,然后再被细胞摄入。吸附内吞有一定的特异性。
3.受体介导的内吞作用(receptor mediated endocytosis) 是细胞依靠细胞表面的受体特异性地摄取细胞外蛋白或其他化合物的过程。细胞表面的受体具有高度特异性,与相应配体(被内吞的分子)结合形成复合物,继而此部分质膜凹陷形成有被小窝,小窝与质膜脱离形成有被小泡,将细胞外物质摄入细胞内。有被小泡进入细胞后,脱去外衣,与胞内体的小囊泡结合形成大的内体,其内容呈酸性,使受体与配体分离。带有受体的部分膜结构芽生、脱落,再与质膜融合,受体又回到质膜,完成受体的再循环。
在内吞过程中,质膜上受体与配体特异结合部位的胞质面(将形成有被小泡的外衣)有一些蛋白附着:①网格蛋白是其中最主要的一种蛋白。它是一种纤维蛋白,与另一种较小的多肽形成了有被小泡外衣的结构单位,即三腿蛋白复合物。三腿蛋白复合物包括三个网格蛋白和三个较小的多肽。由许多三腿蛋白复合物聚合构成五边形或六边形的网格样结构,覆于有被小泡或有被小窝的胞质面。由网格蛋白装配成的外衣提供了牵动质膜的机械力,导致有被小窝的下凹,也有助于捕获膜上的特异受体及与之结合的被转运分子;②调节素是有被小泡中组成外衣的另一类重要的蛋白,它是多亚基的复合物,能识别特异的跨膜蛋白受体,并将其连接至三腿蛋白复合物上,起选择性介导作用。跨膜受体蛋白胞质面肽链尾部,常在一个由四个氨基酸残基构成的区域内高度转折,形成一个内吞信号,由调节素识别它。所以调节
素可介导不同类型受体,使细胞能捕获不同类型的物质。
二外吐作用(exocytosis)
外吐作用又称出胞作用,是一种与内吞作用相反的过程。细胞内物质的分泌,细胞中的病毒、未消化的残渣等分子释放到细胞外都是细胞外吐的过程。
细胞内物质外吐方式
1.固有分泌(constitutive pathway of secretion) 是新合成的分子在高尔基复合体装入转运小泡,随即很快被带到质膜,并持续不断地被细胞分泌出去,它普遍存在于所有细胞内。"SNARE假说"认为在固有分泌中,V-SNARE与t-SNARE相互识别并结合形成7S复合物,这是分泌颗粒与靶膜的特异性结合,接着NSF在SNAP的介导下与7S复合物结合形成20S复合物,此复合物中的SNAP可激活NSF的 ATP酶活性,NSF水解ATP提供能量使20S复合物解聚,随后膜融合自动发生,颗粒内物质分泌到细胞外。
2.受调分泌(regulated pathway of secretion) 是细胞内大分子合成后被贮存在特殊的小泡如分泌颗粒中,只有当细胞接受细胞外信号物质的作用后,引起细胞内一系列生化改变,分泌颗粒才与质膜融合,发生外吐。受调分泌主要存在于特化的分泌细胞,如内、外分泌细胞,神经细胞等。它们能特异性地按需要快速地分泌其产物,如激素、消化酶、神经递质等。 三质膜的循环和运动 在细胞内吞和外吐过程中细胞质膜的面积不断地发生变化。内吞过程中,转运小泡将质膜带人细胞内,如巨噬细胞在吞噬过程中每分钟可以将30%质膜带入细胞;细胞外吐过程中,质膜的面积增大,如一些外分泌细胞分泌消化酶时,可使细胞顶部质膜增大30倍。但细胞的体积和表面积不会因此而增大和缩小,这意味着内吞和外吐的过程是两个相辅相成的过程,即有一定数量的质膜经内吞被减少,就会有相应数量的质膜经外吐过程得以补充,以保持细胞质膜面积的恒定,这就是质膜循环的一种方式。同时,在此循环过程中,质膜也在进行运动,其成分和分布发生了流动,这种流动将有利于细胞功能的执行。
质膜的特化结构包括侧面的特化结构和游离面的特化结构。侧面的特化结构就是指细胞连接,或称细胞间连接,它是细胞相互连接处局部质膜所形成的特化结构,在多细胞动物中普遍存在。游离面的特化结构,如微绒毛、鞭毛、纤毛等,帮助完成细胞的特定活动。 1.紧密连接(tight junction) 又称闭锁小带,存在于上皮细胞近管腔的侧面,多呈带状分布,少数为点状,起着封闭细胞间隙的作用,可防止管腔内物质自由进入细胞间隙。其主要功能为:①通过封闭上皮细胞周围的间隙而阻止。
2022年9月,南京邮电大学团队构建了一种由透明质酸、光敏剂二氢卟吩(Ce6)和甲硝唑(MNZ)组成的纳米试剂(HCM),用于耐甲氧西林金黄色葡萄球菌(MRSA)生物膜感染的治疗。相关成果发表于国际学术期刊《自然·通讯》。
上一篇 细胞内膜系统
下一篇 biomembrane