-
外显子 编辑
 外显子
外显子
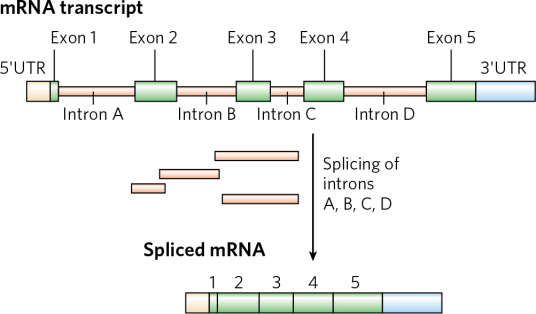
在反式剪接中,不同mRNA的外显子可以被接合在一起。外显子在剪接(Splicing)后仍会被保存下来,并可在蛋白质生物合成过程中被表达为蛋白质。外显子是最后出现在成熟RNA中的基因序列,又称表达序列。既存在于最初的转录产物中,也存在于成熟的RNA分子中的核苷酸序列。术语外显子也指编码相应RNA外显子的DNA中的区域。简言之,外显子就是指真核细胞的基因在表达过程中能编码蛋白质的核苷酸序列。关键概念:比较不同物种的相关基因,发现相应的外显子序列通常是保守的,而内含子序列则很少保守。编码蛋白质的序列通常处于选择压力之下,内含子由于没有选择压力,因此比外显子的进化快得多。通过确定在多种生物中出现的片段来鉴定编码区域,而外显子的保守性可以作为这种鉴定的基础。
人类大部分基因组序列都是被垃圾DNA序列分隔成一段段,给定一个已知的目标蛋白质和基因组序列,在该基因组序列中找出一组子字符串(候选外显子),使得其拼接(剪接)与目标蛋白质最匹配(即去掉垃圾DNA序列)。一个强力方法是寻找基因组序列与目标蛋白质序列间的所有局部相似性。若第一个取自基因组序列的子字符串展示了充分相似性于目标蛋白质,那么这个子字符串可被认为是一个推定的外显子。将推定外显子结构化为基因组序列中的赋权区间,它可用三个参数(l、r、w)来描述,l、r分别是推定的外显子的左边、右边的位置,w为其权重。权重w可反该区间是一个外显子的可能性。链是不重叠赋权区间的任一集合,一个链的总权重是该链中所有区间的权重之和。给定一个推定的外显子集,寻找非重叠的推定的外显子的一个最大集。输入:赋权区间(推定的外显子)集。输出:该集合中区间的最大。
 外显子
外显子
操作步骤及其基本原理是:
⑴基因组DNA经“霰弹法”切成小片段后,克隆在位于“外显子捕捉序列”下游的克隆位点上。
⑵将这些重组载体汇总后感染反转录病毒的专宿包装细胞系(ecotropicretroviralpackagingCELLline)——ψ2细胞系。ψ2细胞提供蛋白质产物使载体(自身不能合成病毒蛋白质)成为反转录病毒在细胞里增殖。当反转录病毒在细胞内转录时,如果插入片段中包含有功能的SA位点,则有可能发生RNA剪接反应而将ⅣS切除。
⑶已剪接和未剪接的病毒RNA都包装在病毒子(virion)中,从细胞培养液中收集后用来感染兼宿反转录病毒包装细胞系(amphotropicretroviralpackagingcellline)PA-317。这使反转录病毒再进行一轮复制,并产生能感染猴肾细胞系COS细胞的高效价病毒原种。这样做是由于上一轮克隆在病毒中的插入片段的剪接效率极低,而在第二轮复制时则大大提高了RNA剪接的机会。
⑷从第二个细胞系PA-317细胞中分离得到的病毒,用来感染组成型产生SV40T(肿瘤)抗原的COS细胞。病毒RNA基因组被反转录,并在载体上的SV40复制起点作用下,以环状DNA附加体形式进行复制。
⑸从COS细胞中回收复制的附加体DNA,经限制性内切酶DpnI酶切后转化细菌。在含卡那霉素(Kn)和5—氯—4—溴—3—吲哚—β—D—半乳糖苷(X-gal)的培养基上挑选转化子。卢—半乳糖苷酶可水解X—gal而生成蓝色产物。因此,不产生β—半乳糖苷酶的转化子菌落则呈白色。
⑹只挑选出白色菌落作进一步研究的材料。白色菌落的生成可以有二种原因。一是由于基因发生突变,使夕—半乳糖苷酶失去活性;二是由于在反转录病毒生活周期的RNA时期中发生了剪接反应,从而丢失了α,β—半乳糖苷酶基因。
⑺如果是基因突变,则大多数将是缺失了载体中的“外显子捕捉”部分,就可用人的口—珠蛋白基因片段为探针作菌落杂交,很快可得到验证。
⑻如果是真正发生了RNA剪接事件,准确的剪接反应可切除作为标记的ⅣS,使人口—珠蛋白基因的第1外显子与落入了捕捉陷阱的插入片段中的外显子序列连接,这可直接测定其序列加以证明。从捕捉到的外显子出发,就可进一步用作探针去从基因组基因文库或cDNA文库中分离出基因。
应用聚合酶链反应-单链构象多态性(polymerasechainreaction-singlestrandconformationpolymorphsim,PCR-SSCP)及DNA直接测序技术检测68例SAD患者和65名正常老年人的早老素-1基因第5外显子。
 外显子
外显子
多数早发性家族性阿尔茨海默氏病(familialalzheimer'sdisease,FAD)与14号染色体上的早老素-1(presenilin-1,PS-1)基因突变有关,少数早发性FAD与PS-2基因突变有关。早老素基因与散发性Alzheimer病(sporadicalzheimer'sdisease,SAD)的关系,目前国内外研究较少,为探讨PS-1基因突变在SAD发病机理中的作用,我们用聚合酶链反应-单链构象多态性(polymerasechainreaction-singlestrandconformationpolymorphism,PCR-SSCP)、DNA直接测序技术检测了68例SAD患者及65名正常老年人PS-1基因第5外显子,发现在SAD患者中也存在PS-1基因第5外显子突。
 外显子
外显子
AdrianKrainerandLucaCartegni为了在外显子跳跃突变中恢复正常的外显子,将一个合成的RS(精氨酸-丝氨酸)结构域融合到能够与特异性外显子结合的反义核苷酸片段上。他们首先检测了ESSENCE方法对乳腺癌基因1(BRCA1)突变的作用,BRCA1在外显子18上的ESE突变会导致外显子跳跃。他们发现将ESSENCE化合物与能够与外显子18反义结合的片段融合作用后在体外能够恢复正常的剪切,其中反义片段和RS片段都是必需的。
研究者接着对另一种外显子跳跃突变导致的疾病模型进行研究。神经退行性疾病脊髓肌肉萎缩由运动神经元存活基因1(SMN1)的功能性拷贝缺失导致。SMN1的缺失可以有SMN2的功能所补偿,但SMN2的外显子7的单核苷酸突变会导致整个外显子缺失。研究者发现用ESSENCE方法也能在体外恢复SMN2突变缺失的外显子7。这个新方法虽然还需要体内的许多研究数据,但显然对相关遗传疾病的治疗确实带来的曙光。
许多基因中遗传上的“无义”片段——即内含子,会妨碍基因指导蛋白质的合成。现在,一篇发表于3月11日期的《自然遗传学》杂志上的文章提出了基因识别这些内含子的新机制。
 外显子
外显子
为进一步验证实验结果,Baralle的研究小组将含有A-T病人中缺失的这个碱基对,共由12碱基对组成的健康基因序列插入到一个完全不同基因的外显子中。令人惊异的是,剪接机制开始把整个外显子当作内含子处理,将该外显子剪切掉。Baralle得出结论:实验结果表明,这个序列行使作用时,能够帮助剪接酶识别内含子;缺失时(如在A-T病人中),就会错移到外显子中去。Baralle猜测,这个序列可能也控制着其它基因的剪接,对引起包括癌症在内的各种疾病有一定作用。
这项工作标志着一次“重大发现”,纽约城哥伦比亚大学的JimManley评价道。Manley认为内含子中间的这段关键序列也许是帮助剪接机制识别和除去内含子的“踏脚石”。在整个基因组中,可能存在“几十个类似机制”来确定内含子的组成