-
原核细胞 编辑
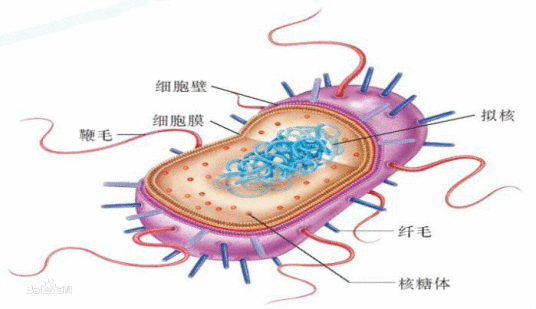
原核细胞(prokaryotic cell)没有核膜,遗传物质集中在一个没有明确界限的低电子密度区。DNA为裸露的环状分子,通常没有结合蛋白,环的直径约为2.5纳米,周长约几十纳米。没有恒定的内膜系统,核糖体为70S型,原核细胞构成的生物称为原核生物(prokaryote),均为单细胞生物,通常称为细菌(bacterium)。
原核细胞( procaryotic/prokaryotic cell )指没有核膜且不进行有丝分裂、减数分裂、无丝分裂的细胞。这种细胞不发生原生质流动,观察不到变形虫样运动。鞭毛(flagellum)呈单一的结构。光合作用、氧化磷酸化在细胞膜进行,没有叶绿体(chloroplast)、线粒体
细胞壁 厚度因细菌不同而异,一般为15-30纳米。主要成分是肽聚糖,由N-乙酰葡糖胺和N-乙酰胞壁酸构成双糖单元,以β(1-4)糖苷键连接成大分子。N-乙酰胞壁酸分子上有四肽侧链,相邻聚糖纤维之间的短肽通过肽桥(革兰氏阳性菌)或肽键(革兰氏阴性菌)桥接起来,形成了肽聚糖片层,像胶合板一样,粘合成多层。肽聚糖中的多糖链在各物种中都一样,而横向短肽链却有物种间的差异。革兰氏阳性菌细胞壁厚约2080纳米,有15-50层肽聚糖片层,每层厚1纳米,含20-40%的磷壁酸(teichoic acid),有的还含有少量蛋白质。革兰氏阴性菌细胞壁厚约10纳米,仅2-3层肽聚糖,其他成分较为复杂,由外向内依次为脂多糖、细菌外膜和脂蛋白。此外,外膜与细胞之间还有间隙。
肽聚糖是革兰阳性菌细胞壁的主要成分,凡能破坏肽聚糖结构或抑制其合成的物质,都有抑菌或杀菌作用。如溶菌酶是N-乙酰胞壁酸酶,青霉素抑制转肽酶的活性和肽桥的形成。
细菌细胞壁的功能包括:保持细胞外形;抑制机械和渗透损伤(革兰氏阳性菌的细胞壁能耐受20kg/c㎡的压力);介导细胞间相互作用(侵入宿主);防止大分子入侵;协助细胞运动和分裂。
脱壁的细胞称为细菌原生质体(baCTerial protoplast)或球状体(spheroplast),因脱壁不完全,脱壁后的细菌原生质体,生存和活动能力大大降低。
细胞膜
是典型的单位膜结构,厚约8-10纳米,外侧紧贴细胞壁,某些革兰氏阴性菌还具有细胞外膜。细胞膜有多方面的重要功能,它与细胞的物质交换,细胞识别、分泌、排泄、免疫等都有密切的关系。通常不形成内膜系统,除核糖体外,没有其它类似真核细胞的细胞器,呼吸和光合作用的电子传递链位于细胞膜上。某些行光合作用的原核生物(蓝细菌和紫细菌),质膜内褶形成结合有色素的内膜,与捕光反应有关。某些革兰氏阳性细菌质膜内褶形成小管状结构,称为中膜体(mesosome)或间体,中膜体扩大了细胞膜的表面积,提高了代谢效率,有拟线粒体(Chondroid)之称,此外还可能与DNA的复制有关。
细胞质与核质体
细菌和其它原核生物一样,没有核膜,DNA集中在细胞质中的低电子密度区,称核区或核质体(nuclear body)。细菌一般具有1-4个核质体,多的可达20余个。核质体是环状的双链DNA分子,所含的遗传信息量可编码2000-3000种蛋白质,空间构建十分精简,没有内含子。由于没有核膜,因此DNA的复制、RNA的转录与蛋白质的合成可同时进行,而不像真核细胞那样在生化反应在时间和空间上是严格分隔开来的。
每个细菌细胞约含5000-50000个核糖体,部分附着在细胞膜内侧,大部分游离于细胞质中。细菌核糖体的沉降系数为70S,由大亚单位(50S)与小亚单位(30S)组成,大亚单位含有23SrRNA与30多种蛋白质,小亚单位含有16SrRNA与20多种蛋白质。30S的小亚单位对四环素与链霉素很敏感,50S的大亚单位对红霉素与氯霉素很敏感。
细菌核区DNA以外的,可进行自主复制的遗传因子,称为质粒(plasmid)。质粒是裸露的环状双链DNA分子,所含遗传信息量为2-200个基因,能进行自我复制,有时能整合到核DNA中去。质粒DNA在遗传工程研究中很重要,常用作基因重组与基因转移的载体。
胞质颗粒是细胞质中的颗粒,起暂时贮存营养物质的作用,包括多糖、脂类、多磷酸盐等。
拟核
拟核(nucleoid)存在于原核生物,是没有由核膜包被的细胞核,只有一个位于形状不规则且边界不明显区域的环形DNA分子,核区没有强的Feulgen阳性反应,但是人们沿用了真核细胞的染色体概念,也把细胞的核区DNA称为染色体,实际上它没有真正的染色体结构 。内含遗传物质。里面的核酸为双股螺旋形式的环状DNA,且同时具有多个相同的复制品。
其他结构
许多细菌的最外表还覆盖着一层多糖类物质,其中边界明显的称为荚膜,如肺炎球菌,边界不明显的称为粘液层(slime layer),如葡萄球菌。荚膜对细菌的生存具有重要意义,细菌不仅可利用荚膜抵御不良环境;保护自身不受白细胞吞噬;而且能有选择地粘附到特定细胞的表面上,表现出对靶细胞的专一攻击能力。例如,伤寒沙门杆菌能专一性地侵犯肠道淋巴组织。细菌荚膜的纤丝还能把细菌分泌的消化酶贮存起来,以备攻击靶细胞之需。
鞭毛是某些细菌的运动器官,由一种称为鞭毛蛋白(flagellin)的弹性蛋白构成,结构上不同于真核生物的鞭毛。细菌可以通过调整鞭毛旋转的方向(顺时针和逆时针)来改变运动状态。
菌毛是菌体表面极其细的蛋白纤维,须用电镜观察,特点是:细、短、直、硬、多。菌毛与细菌运动无关,根据形态、结构和功能,可分为普通菌毛和性菌毛两类。前者与细菌吸附和侵染宿主有关,后者为中空管子,与传递遗传物质有关。
原核生物的基因结构多数以操纵子形式存在,即完成同类功能的多个基因聚集在一起,处于同一个启动子的调控之下,下游同时具有一个终止子。两个基因之间存在长度不等的间隔序列,如与乳糖代谢有关酶的基因。在距转录起始点-35和-10(转录起始点上游的核苷酸序列为“-”,下游的核苷酸序列为“+”)附近的序列都有RNA聚合酶识别的信号。RNA聚合酶先与-35附近的序列(称为Pribnow框)结合,然后才与-10附近的序列(称为Sextama框)结合。RNA聚合酶一旦与-10附近序列结合,就立即从识别位点上脱离下来,DNA双链解开,转录开始。除启动子外,往往还有一些调控转录的其他因子,如调节基因和操纵基因。
原核生物基因转录终止之前同样有一段回文序列结构,称为终止子,它的特殊的碱基排列顺序能够阻碍RNA聚合酶的移动,并使其从DNA模板链上脱离下来。
细菌以一分为二的方式繁殖,某些细菌处于不利的环境,或耗尽营养时,形成内生孢子,又称芽孢,是对不良环境有强抵抗力的休眠体,由于芽胞在细菌细胞内形成,故常称为内生孢子。
芽孢的生命力非常顽强,有些湖底沉积土中的芽孢杆菌经500-1000年后仍有活力,肉毒梭菌的芽孢在pH7.0时能耐受100℃煮沸5-9.5小时。芽孢由内及外有以下几部分组成:
1.芽孢原生质(spore protoplast):核心(core)含浓缩的原生质。
2.内膜(inner membrane):由原来繁殖型细菌的细胞膜形成,包围芽孢原生质。
3.芽孢壁(spore wall):由繁殖型细菌的肽聚糖组成,包围内膜。发芽后成为细菌的细胞壁。
4.皮质(cortex):是芽孢包膜中最厚的一层,由肽聚糖组成,但结构不同于细胞壁的肽聚糖,交联少,多糖支架中为胞壁酐而不是胞壁酸,四肽侧链由L-Ala组成。
5.外膜(outer membrane):也是由细菌细胞膜形成的。
6.外壳qiao(coat):芽孢外壳,质地坚韧致密,由类角蛋白(keratinlike protein)组成,含有大量二硫健,具有疏水性特征。
传统分类法根据生物的营养方式、运动能力和细胞结构的特点,把生物划分为动物界和植物界。植物细胞的主要特征是具有硬的细胞壁和进行光合作用的叶绿体。按传统分类系统,虽然大多数生物种容易归类,可是对某些生物来说却遇到了分类上的困难,例如眼虫(Euglena)是一种单细胞生物,含有叶绿体,却不具有细胞壁;细菌和真菌则有细胞壁而无叶绿体;支原体既无叶绿体也无细胞壁,古细菌既有原核生物的特征也具有真核生物的特征。这些生物按照传统分类法进行分类显然就要遇到困难。
1977年C. Woese根据对16SrRNA核苷酸顺序的同源性比较,提出将生命划分为三界,即:真细菌(Eubacteria)、真核生物Eucaryotes、古细菌(Archaes)。1996年Bult领导的研究小组在Science上发表了詹氏甲烷球菌(MethaNOcoccus jannaschii)的全基因组序列,进一步证明它既不是典型的细菌也不是典型的真核生物,而是介于两者之间的生命体,即生命的第三形式。
原核生物(prokaryote)是以原核细胞构成的,均为单细胞生物,通常称为细菌(bacterium)。
根据外表特征,可把原核生物粗分为“三菌三体”6种类型,即细菌(狭义的)、放线菌、蓝细菌、支原体、立克次氏体和衣原体。
细菌
 蓝藻
蓝藻
支原体
 支原体
支原体
支原体基因组为一环状双链DNA,分子量小(仅有大肠杆菌的五分之一),合成与代谢很有限。肺炎支原体的一端有一种特殊的末端结构(terminal structure),能使支原体粘附于呼吸道粘膜上皮细胞表面,与致病性有关。
衣原体和立克次氏体
衣原体(Chlamydia)很小,直径200-500纳米,能通过细菌滤膜。立克次氏体(Rickettsia)略大,大多不能通过滤菌膜。它们都有DNA和RNA,有革兰氏阴性细菌特征的含肽聚糖的细胞壁,但酶系统不完全,必须在寄主细胞内生活,有摄能寄生物(energe parasite)之称。
砂眼是衣原体引起的,由于能形成包含体,起初被认为是大型病毒,1956年,中国著名微生物学家汤飞凡及其助手张晓楼等人首次分离到沙眼的病原体。
 立克次氏体
立克次氏体
美国医生H.T.Richetts 1909年首次发现它是落基山斑疹伤寒的病原体,并于1910年牺牲于此病,故后人称这类病原体为立克次氏体。与衣原体的不同处在于其细胞较大,无滤过性,合成能力较强,且不形成包涵体。