-
糖苷酶 编辑
糖苷酶即糖苷水解酶(Glycoside hydrolases,GH,EC3.2.1),是一类水解糖苷键(glycosidic bonds)的酶,在生物体糖和糖缀合物的水解与合成过程中扮演着重要角色。糖苷酶在催化糖苷反应时,如果水分子的氧原子进攻受体葡萄糖上的异头碳,即发生水解反应,但如果是葡萄糖羟基上的氧原子进攻受体葡萄糖上的异头碳,即发生转糖基反应。
糖苷酶几乎存在于所有的生物体中,是一类以内切或外切方式水解各种含糖化合物(包括单糖苷、寡糖、多糖、皂甙和糖蛋白等)中的糖苷键,生成单糖、寡糖或糖复合物的酶。糖苷酶在寡糖合成、烷基糖苷和芳香基糖苷的合成、氨基酸和多肽的糖基化以及抗生素的糖基化方面发挥了重要作用。
随着采用广泛的生物材料进行酶的提纯和性质研究的进展,已知在生物体中存在着许多种类的糖苷酶,利用这些酶的特异的水解作用,已作为研究糖苷、寡糖、多糖的必不可少的试剂。另外对这些酶的分布、存在形态、形式结构、遗传缺陷的研究,具体地阐明了这些酶在生物体内的正常代谢中所起的作用。糖苷酶一般对糖苷配基的特异性比对糖残基的特异性差,因此有许多仍未能确定其“真正的天然底物”到底是什么。常根据糖残基的性质(包括糖苷键的性质)来进行命名和分类的(如α-葡糖苷酶、β-半乳糖苷酶),另也常使用历史上最初发现时来自底物名的惯用名(如蔗糖酶=β-果糖苷酶,麦芽糖酶=α-葡糖苷酶)。
糖苷酶(EC 3.2.1)隶属于糖基酶(EC 3.2),是水解酶类(EC 3)中的一大类酶。根据不同的分类标准,可以分成不同的类型。
根据底物分类
在糖苷酶家族中,根据水解的底物不同,又可具体分为195种不同类型的酶,每种均有自己的酶学编号(EC 3.2.1.1——EC 3.2.1.195),例如α-淀粉酶(EC 3.2.1.1)与α-葡萄糖苷酶(EC 3.2.1.20)。 此分类方法中,分类对象只有糖苷酶,每一种编号对应的糖苷酶成员之间的结构可能会有巨大的差异。
根据结构相似性分类
根据进化学分析,许多水解不同底物的糖苷酶是由同一祖先演化而来,因而具有较高的结构与功能相似性。根据结构的差异,可将糖苷酶具体分为135个家族(Glycoside Hydrolase family)即GH1——GH135,每个家族内部的成员,彼此具有很高的结构相似度,根据其催化活性中心结构域的结构,又可将这些家族不完全归纳为14个族(Clan)即GH-A —— GH-N。 此分类方法中,分类对象除了糖苷酶还包括少量其他种类的酶,同一家族中的成员虽然水解的底物可能不同,但都具有高度相似的结构,彼此间的亲缘关系更为接近。
根据催化作用机制分类
糖苷酶根据催化作用机制的不同分为两类:构型翻转酶( invERTing en-zymes)和构型保持酶(retaining enzymes)。构型保持酶在催化糖苷键水解的同时,还具有转糖基活性,即糖苷键合成活性,该性质使其成为糖类合成的重要工具。
1、α-甘露糖苷酶。在N-糖基化过程中所涉及的甘露糖苷酶,依据它们所催化水解键位的不同主要可分为α-甘露糖苷酶和β-甘露糖苷酶,它们分别参与甘露糖α-1, 2、α-1, 3、α-1, 6和β-1, 4糖苷键的水解过程。迄今为止,已克隆的α-甘露糖苷酶cDNA 20多种,其中来自人类的6种。α-甘露糖苷酶主要存在于内质网、高尔基体、溶酶体和胞浆其它细胞器中,功能是剪接寡糖结构末端不同连接的甘露糖残基,形成高甘露糖型、复杂型、杂合型的N-寡糖。依据底物特异性、酶分子量大小、保守序列的结构域及对生物碱抑制作用的敏感性,Moremen于1994年把已经发现的α-甘露糖苷酶分成Ⅰ类和Ⅱ类两大类。
2、阿拉伯糖苷酶。现已提纯的阿拉伯糖苷酶,根据其氨基酸序列,基本上属于糖基水解酶3、10、43、51、54、64这6个家族。根据底物专一性的异同,阿拉伯糖苷酶又可分为两类:阿拉伯糖苷酶A一般降解末端的阿拉伯糖基,阿拉伯糖苷酶B既能降解末端的阿拉伯糖基,也能切阿拉伯糖基的侧枝。在细菌和真菌中都已有阿拉伯糖苷酶被提纯和定性。阿拉伯糖苷酶可分布在胞内、胞外和胞膜上,有的是单聚体,而有的形成二聚体或多聚体,如从Butyrivibrio fibrisolvensGS113提纯的阿拉伯糖苷酶就是个八聚体。而且有些阿拉伯糖苷酶是由不同的亚基聚集而成的,如Bacillus stearothermophilus产生的阿拉伯糖苷酶含有两个不同的亚基。
3、β-木糖苷酶。β-木糖苷酶在自然界中分布广泛,现已从细菌、放线菌和真菌(包括酵母)等微生物和高等植物中分离得到。β-木糖苷酶是一种外切酶,主要催化水解木糖苷,并以外切方式从非还原性末端水解木二糖及木二糖以上的低聚木糖,水解产物为木糖。木聚糖类半纤维素酶解时一般由木聚糖酶从主链内部先作用于长链木聚糖的糖苷键上,将木聚糖随机切成不同链长的低聚木糖,再由β-木糖苷酶作用于低聚木糖的末端,将这些短链低聚木糖降解成木糖。β-木糖苷酶还可以作用于萜类、甾体等甙元与木糖形成的糖苷键,释放出甙元。
4、壳三糖苷酶。壳三糖苷酶(Chitotriosidase, CT)是一种功能性几丁质酶(E. C. 3. 2. 1. 14)。几丁质酶是一类可以水解几丁质的蛋白质,存在于原核生物和真核生物的多种物种体内,因其与物种受到外来侵袭后的防御反应以及信号转导等有密切关系,所以在抵抗外来病原体,特别是植物抗真菌病的基因工程中应用广泛。人类曾被认为是没有也不能降解几丁质成分的,因此长期以来几丁质酶均被作为化疗药物,如抗真菌药物、抗疟疾药物、杀虫剂等的靶标。但是最近,Boot等发现了人体内也存在几丁质酶,将其称为壳三糖苷酶。核酸序列分析发现,壳三糖苷酶与其他物种的几丁质酶家族成员具有高度同源性。
5、硫代糖苷酶。微生物糖苷酶来源广泛,种类繁多,有些糖苷酶除具有水解活性外,还具有转基活性,该性质使其成为糖类合成的重要工具,被用于大规模合成多种O-糖苷。近三年研究发现,微生物糖苷酶的一类新型突变酶即硫代糖苷酶(thioglycoligases)能催化硫代糖苷(thioglycosides)的合成,这一发现引起了科学家的极大兴趣。硫代糖苷是O-糖苷类似物,糖单位组成和空间结构与O-糖苷类似,不同之处仅在于糖苷键通过硫原子起连接作用,不易被糖苷酶水解,具有重要的研究价值:由于化学水解和酶解速率低,可以解决O-糖苷易被内源糖苷酶水解的问题,从而作为O-糖苷替代品,应用于药物疗法;作为糖苷酶的竞争性抑制剂,与糖苷酶形成稳定的复合物用于X-射线晶体结构分析,研究糖苷酶特异性和作用机制,探索其突变或缺陷引起人类疾病的分子机理;用于制备亲和树脂纯化糖苷酶蛋白;作为非降解性配体用于凝集素研究等。由于硫代糖苷在生物技术和制药业方面的潜在价值越来越受到关注,相应地,其大量获得也成为当今研究的热点。
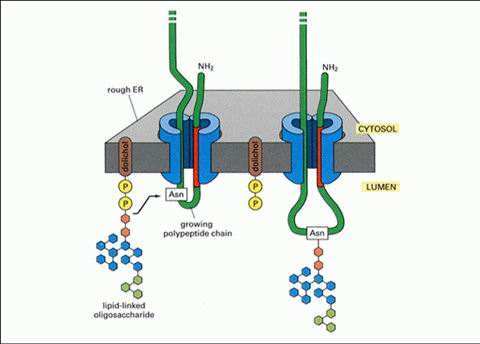
N-糖基化是在内质网上由糖基转移酶催化,在内分泌蛋白和膜结合蛋白的天冬酰氨残基的氨基上结合寡糖的过程,即在粗面内质网的核糖体上合成蛋白肽链的同时,一旦形成天冬氨酸-Xaa-色氨酸-/丝氨酸(Asn-Xaa-Ser/Thr, Xaa为除脯氨酸外的所有氨基酸残基)三联序列子密码,即糖基化位点,才有可能开始糖基化,随着肽链进入内质网腔,N-糖链可被位于内质网腔膜结构上的加工酶修剪、加工成高甘露糖型,再进入高尔基体,继续被膜结构上的酶加工合成。少数情况下,天冬氨酸-Xaa-半光氨酸(Asn-Xaa-Cys)序列也作为糖基化位点。N-糖基化的过程中涉及多种酶的参与,主要是糖酰基转移酶和糖苷酶两大类,前者负责将活性供体(通常是NDP-糖)的单糖部分转移至接纳体物质如糖、蛋白质和脂等分子上;后者的催化活性是修剪糖蛋白糖链上的各种糖基,二者共同完成N-糖基化。