-
趋化因子 编辑
 趋化因子
趋化因子
趋化因子蛋白的共同结构特征包括,分子量小(约8-10 kDa),有四个位置保守的半胱氨酸残基以保证其三级结构。
一部分趋化因子被认为是促炎性细胞因子,可以在免疫应答过程中诱导免疫系统的细胞进入感染部位。而有些趋化因子被认为维持机体自我调节,在正常的组织维持或发育过程中控制细胞的迁徙。在所有的脊椎动物、一些病毒和一些细菌中有趋化因子存在,但在其他无脊椎动物中尚未发现。
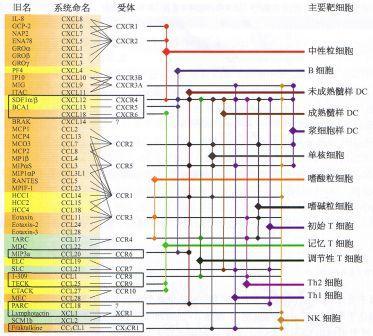
趋化因子被分为四个主要亚家族:CXC、CC、CX3C和XC。所有这些蛋白都通过与G蛋白连接的跨膜受体(称为趋化因子受体)相互作用来发挥其生物学效应这些蛋白质结合到趋化因子受体而起作用,趋化因子受体是G蛋白偶连的跨膜受体,选择性地表达在靶细胞表面。
 趋化因子有四个位置保守的半胱氨酸残基以保证其三级结构
趋化因子有四个位置保守的半胱氨酸残基以保证其三级结构
所有趋化因子都很小,分子量在8到10 kDa之间。它们之间大约有20-50%是相同的;也就是说,它们具有相同的基因序列和氨基酸序列同源性。它们也都拥有保守的氨基酸,这些氨基酸对形成它们的三维或三级结构很重要,例如在大多数情况下四个半胱氨酸相互作用,形成一个希腊钥匙形状,这是趋化因子的一个特征。分子内二硫键通常连接第一个到第三个半胱氨酸残基,第二个到第四个半胱氨酸残基,它们在趋化因子的蛋白序列中出现时编号。典型的趋化因子蛋白是以肽前体的形式产生的,在其从细胞分泌的过程中,从分子的活性(成熟)部分分裂出大约20个氨基酸的信号肽。在趋化因子中,前两个半胱氨酸靠近成熟蛋白的N端,第三个半胱氨酸位于分子中心,第四个靠近C端。在前两个半胱氨酸之后有一个大约10个氨基酸的环,称为N环。
趋化因子的主要作用是诱导细胞定向迁移,被趋化因子吸引的细胞沿着趋化因子浓度增加的信号向趋化因子源处的迁徙。有些趋化因子在免疫监视过程中控制免疫细胞趋化,如诱导淋巴细胞到淋巴结,这些淋巴结中的趋化因子通过与这些组织中的抗原提呈细胞相互作用而监视病原体的入侵。这些被称为稳态趋化因子,在不需要刺激源细胞的情况下产生和分泌。有些趋化因子在发育中起作用;他们能刺激新血管形成(血管新生);引导细胞进入组织,为细胞的成熟提供特定的信号。其他趋化因子在炎症反应中起关键作用,可以因应对细菌感染、病毒感染而由多种细胞释放;也可以因非感染性的刺激如二氧化硅吸入,尿路结石等而释放。趋化因子的释放通常由炎症细胞因子如白细胞介素1 (IL-1) 刺激引起。炎性趋化因子的主要作用是作为白细胞的趋化剂,从血液中吸引单核细胞、中性粒细胞和其他效应细胞到感染或组织损伤的部位。某些炎性趋化因子可激活细胞来启动免疫反应或促进伤口愈合。它们由许多不同的细胞类型释放,并对天然免疫系统和适应性免疫系统的细胞起引导作用。
趋化因子的主要功能是在炎症和体内平衡过程中管理白细胞向各自位置的迁移(归巢)。
基础归巢作用:在胸腺和淋巴组织中产生的基础的稳态趋化因子。趋化因子CCL19和CCL21(在淋巴结和淋巴管内皮细胞中表达)及其受体CCR7(在细胞中注定要归巢到这些器官的细胞中表达)是它们在归巢中的稳态功能的最好例证。利用这些配体可使抗原呈递细胞(APC)在适应性免疫应答过程中转移至淋巴结。其他的稳态趋化因子受体包括:CCR9、CCR10和CXCR5,它们作为细胞地址的一部分对于组织特异性的白细胞归巢非常重要。CCR9支持白细胞向肠内迁移,CCR10支持皮肤迁移,CXCR5支持b细胞向淋巴结滤泡迁移。骨髓中生成的CXCL12 (SDF-1)促进了骨髓微环境中B祖细胞的增殖
炎症归巢作用:炎症趋化因子在感染或损伤过程中产生高浓度,并决定炎症性白细胞向受损区域的迁移。典型的炎性趋化因子包括:CCL2、CCL3和CCL5、CXCL1、CXCL2和CXCL8。一个典型的例子是CXCL-8,它是中性粒细胞的趋化剂。与稳态趋化因子受体相比,结合受体和炎症趋化因子存在明显的混杂。这常常使受体特异性治疗的研究复杂化。
结构分类
趋化性细胞因子根据其氨基端(N端)半胱氨酸的排列方式,可分为CXC、CC、XC和CX3C四个亚族:
CC趋化因子亚族:CCL1、CCL2、CCL3、CCL4、CCL5、CCL6、CCL7、CCL8、CCL9、CCL10、CCL11、CCL12、CCL13、CCL14、CCL15、CCL16、CCL17、CCL18、CCL19、CCL20、CCL21、CCL22、CCL23、CCL24、CCL25、CCL26、CCL27、CCL28
CXC趋化因子亚族:CXCL1、CXCL2、CXCL3、CXCL4、CXCL5、CXCL6、CXCL7、CXCL8、CXCL9、CXCL10、CXCL11、CXCL12、CXCL13、CXCL14、CXCL15、CXCL16、CXCL17
XC趋化因子亚族:XCL1、XCL2
CX3C趋化因子亚族:CX3CL1
功能分类
趋化因子根据其功能不同分为两类:
体内平衡趋化因子:在某些组织中产生,负责基础白细胞迁移。包括:CCL14、CCL19、CCL20、CCL21、CCL25、CCL27、CXCL12和CXCL13。(这种分类并不严格,例如:CCL20也可以作为促炎趋化因子)
促炎趋化因子:在病理条件下形成的(在促炎刺激下,如IL-1、TNF-α、LPS或病毒),并积极参与炎症反应,吸引免疫细胞到炎症部位。例如:CXCL-8、CCL2、CCL3、CCL4、CCL5、CCL11、CXCL10。
作用细胞分类
趋化因子根据其趋化作用的细胞类型不同,可以分为如下几类:
单核/巨噬细胞趋化因子:吸引单核/巨噬细胞到炎症部位的关键趋化因子包括:CCL2、CCL3、CCL5、CCL7、CCL8、CCL13、CCL17和CCL22。
T淋巴细胞趋化因子:参与T淋巴细胞募集到炎症部位的四个关键趋化因子是:CCL2、CCL1、CCL22和CCL17。此外,T细胞激活后诱导CXCR3表达,活化的T细胞被炎症部位吸引,在炎症部位分泌IFN-γ诱导的趋化因子CXCL9、CXCL10和CXCL11。
肥大细胞趋化因子:表面表达多种趋化因子受体:CCR1、CCR2、CCR3、CCR4、CCR5、CXCR2、CXCR4。这些受体CCL2和CCL5的配体在肺肥大细胞募集和活化中起重要作用。也有证据表明CXCL8可能抑制肥大细胞。
嗜酸性粒细胞趋化因子:嗜酸性粒细胞向各种组织的迁移涉及CC家族的几种趋化因子:CCL11、CCL24、CCL26、CCL5、CCL7、CCL13和CCL3。趋化因子CCL11 (eotaxin)和CCL5 (rantes)通过嗜酸性粒细胞表面的一个特定受体CCR3发挥作用,而嗜酸性粒细胞在最初募集到病变中发挥重要作用。
中性粒细胞趋化因子:主要由CXC趋化因子调节。例如,CXCL8 (IL-8)是中性粒细胞的趋化剂,并激活其代谢和脱颗粒。
趋化因子受体是在白细胞表面发现的含有7个跨膜结构域的G蛋白偶联受体。迄今为止,大约已经鉴定出19种不同的趋化因子受体,根据它们结合的趋化因子的类型,它们被分为四个家族:与CXC趋化因子结合的CXCR,与CC趋化因子结合的CCR,与唯一CX3C趋化因子(CX3CL1)结合的CX3CR1,与两个XC趋化因子(XCL1和XCL2)结合的XCR1。它们有许多共同的结构特征:它们的大小相似(约有350个氨基酸),有一个短的酸性N端,7个螺旋跨膜结构域,3个胞内和3个胞外亲水环,以及一个胞内含有对受体调节重要的丝氨酸和苏氨酸残基的C端。前两个细胞外环的趋化因子受体,每个都有一个保守的半胱氨酸残基,允许形成一个二硫键之间的这些环。G蛋白偶联到趋化因子受体的C端,使受体激活后胞内信号传导,而趋化因子受体的N端结构域决定了配体结合的特异性。
信号转导
趋化因子受体与G蛋白结合后传递细胞信号。G蛋白被趋化因子受体激活,导致随后一种被称为磷脂酶C (PLC)的酶激活。PLC将一种称为磷脂酰肌醇(4,5)-二磷酸(PIP2)的分子分裂成两个第二信使分子,称为肌醇三磷酸(IP3)和二酰基甘油(DAG),从而触发细胞内信号事件;DAG激活另一种叫做蛋白激酶C (PKC)的酶,而IP3则触发细胞内钙的释放。这些事件促进许多信号级联反应(如MAP激酶途径),产生趋化、脱颗粒、释放超氧阴离子以及细胞内包涵趋化因子受体的粘着分子(称为整合素)的活性变化。