-
种内关系 编辑
中文名:种内关系
外文名:intraspecificrelationship
动物
大量实验研究已揭示出密度效应的各种机制。例如,当把一对果蝇置于一个瓶中,并供应一定量的食物时,起初其后代数量会迅速增加,很快就会达到一定限度。但是,当把很多果绳置于同样的瓶中时,每对果蝇所产出的后代数量将与瓶中采蝇的密度成反比。这种效应是由于幼虫竞争食物引起的,密度越大竞争所引起的死亡率越高,成虫的寿命也会下降,引起成虫寿命下降的密度比引起幼虫存活率下降的密度高。密度制约因素对未成年期的有害影响往往比成年期大。陈艳等(1998)对美洲斑潜蝇实验种群密度效应研究结果表明,卵期不存在密度效应,叶片上的卵量及分布与潜蝇成虫数量的多少有关;幼虫期存在密度效应,1龄幼虫对幼虫期存活率、预蛹重、成虫寿命、繁殖等有显著的影响。密度超过0.8头/cm2时,由于空间不足、营养不足、排泄物及脱皮物增多,生态环境恶化,个体发育不良。同时,营养不足又使幼虫间“自残”(cannibalism)。
植物
植物种群内个体间的竞争,主要表现为个体间的密度效应,反映在个体产量和死亡率上。因为植物不能像动物那样逃避密集和环境不良的情况,其表现只是在良好情况下可能枝繁叶茂,而高密度下可能枝叶少、构件数少。植物密度效应有两个基本的规律。
2.1.最后产置恒值法则
最后产量恒值法则(law of constant final yield)是指在一定范围内,当条件相同时,不管一个种群的密度如何,最后产量差不多总是一样的。澳大利亚生态学家Donald(1951)对三叶草的密度与产量关系做了一系列研究后,证实了这个法则。
2.2. -3/2 方自疏法则
生长于较高密度种群内的植物,由于密度的抑制作用,种群内个体会逐渐死亡,种群数目逐渐减少,直至达到平衡。这种种群的生长动态现象就称为自然稀疏或者自疏(self-thinning)。 自疏现象普遍存在于自然和人工植物种群中, 与农业、林业以及牧草生产有着密切的联系。由密度导致的植物种群的自然稀疏现象,是植物种群生态学研究的热点,对于植物自然稀疏规律(种群密度和个体平均质量的幂律关系)的机理,一直有不同的理论解释。
由-3/2方自疏法可知,在一个空间拥挤、年龄均等、构成单一的植物种群中,随着个体植株生物量的增长,其种群密度将相应减少,且在双对数坐标系里,lgW-lgd的点运动轨迹不会超越一条斜率为-3/2的直线上限,而与植物的生活型、年龄大小、土壤状况及其他栖息地条件等因素无关。-3/2方自疏法则描述了棺物种群在自疏过程中,存活个体的平均质量(W)增加与逐渐减小的密度(d)之间动态关系的经验规律,此规律为植物种群所特有,广泛适用于大小悬殊、形态各异的各种植物。但也有许多关于植物自疏过程的研究结果表明,在自疏过程中平均个体生物量和密度之间虽为指数关系,其数值却与-3/2有较大的偏离,因此对于-3/2方自疏法的普适性产生了争论,争论的焦点主要在于自疏指数的值是等于、大于或是小于-3/2,也就是说植物种群是沿怎样的自然稀疏的轨迹线-3/2上限边界线历程自疏的。
植物性别
植物性别的产生可以追溯到最原始的原核生物细菌的拟有性生殖,例如大肠杆菌进行拟有性生殖时,具有性绒毛的菌体细胞通过性绒毛把遗传物质转移到另一菌体细胞的菌体细胞可看成是雄性配子或个体,即产生了性别的分化。在真核的单细胞植物中,两个营养细胞直接转化为两性配子并以细胞接触处融合,进而完成细胞质和细胞核融合形成合子,其中一个为雄配子,另一个为雌配子,这种繁殖方式称为同配生殖(isogamic reproduCTion)。随着植物体结构进化和发展,产生了异配生殖(anisogamic reproduction )和卵式生殖(oogamicreproduction),同时植物个体也发生了性别差异,产生了雌、雄配子体。在低等植物中,配子体能独立生活,而在髙等植物中,配子体退化成结构简单并寄生在孢子体上的性器宫。植物的性别差异源于有性生殖,大多数生物学家认为在有性生殖过程中发生了遗传物质的重组,由此促进了遗传变异,使生物的进化速率加快,保证了物种能适应复杂多变的环境,并不断地繁衍后代使之延续下去。植物性别具有多态性,例如雄蕊和雌蕊分别单独存在于不同的花中,这种现象就是雌雄异花现象,这样的花就是单性花(unisexual flower)或不具备花 (imperfECT flower)。单性花又分为雄花(staminate)和雌花(pistillate)两种。而雄蕊和雌蕊同时存在于一朵花的现象就是雌雄同花现象,只要雌蕊群和雄蕊群两者都存在且充分发育,这样的花就称两性花。在个体和群体水平上,植物同样存在各种各样的性别类型。最常见的是全部为两性花的植株和群体,该物种为雌雄同花植物。
动物婚配制度
2.1.婚配制度的定义与进化
婚配制度是动物行为生态学的重要研究内容之一。在漫长的进化过程中,动物除了在形态解剖、生理生化及遗传学等方面对环境产生适应性变化之外,也产生了与环境相适应的行为,其中婚配制度、双亲行为、攻击和友好行为等社会行为充分表现了以社群为单位的动物对特定环境的适应性。动物婚配制度的定义需要考虑:①在一个繁殖季里一个个体所获得的配偶数及配偶如何获得;②两性个体是否都具有双亲行为;③两性个体是否形成紧密的配对关系及配对关系维持时间长短。因此,动物婚配制度指的是根据同种群动物个体在一个繁殖季节里获得配偶数多寡、两性个体是否都具有双亲行为、两性个体配对关系(pair-bonding)的紧密程度和持续时间而划分的雌雄婚配分类系统。
由于雌雄两性在婚配中投资不平衡性,一雄多雌制是动物最常见的婚配制度,一雄一雌的单配制则是由原始的一雄多雌的多配制进化而来的。多配制动物生长发育迅速、繁殖潜力高、种群易于扩散,同时可以在资源被消耗尽前迁移到别处建立新的种群,并能利用孤立的生境迅速繁殖。而单配制的种类,一般居住在稳定的环境中,生理成熟和性成熟比较慢,动物能有效地利用资源。单配制动物的胎仔数也较少,具较强的双亲行为,由此确保幼仔从限制性的资源上获得更充分的利益。在高等动物婚配关系中,一般雌性是限制者,雄件常常不易接近雌性,是被限制者。由于这些原因,雄性常常因竞争雌性时发生格斗,或者建立吸引雌性的领域等。
2.2. 影响婚配制度的生态因子
在不同生态因子作用的条件下,动物婚配制度备很大的变异性,许多种类可有几种婚配制度,并且随年龄、季节、地理位置而变化。婚配制度也随种群动态、种群密度的变化而变化。决定动物婚配制度的主要生态因子可能是资源的分布,尤其是食物和营巢地在空间和时间上的分布。从空间上来看,高质量均匀分布的资源布利于产生一雄一雌的单配偶制;相反资源虽然质量较高,但呈斑块状分布的则有利于多配制的产生。
2.3. 婚配制度的类型
按配偶数婚配制度分为单配偶制(moNOgamy)和多配偶制(polygamy),后者又可分一雄多雌制(polygyny)、一雌多雄制(polyandry)和混配制。
(1) 单配偶制。非生物学家往往认为单配偶制是高等动物的特点。实际上动物界内单配偶制是比较少见的,只有鸟类以一雄一雌制较普遍,例如天鹅、丹顶鹤和许多鸣禽。哺乳类中单配偶制倒是例外,如狐、鼬和河狸,其他脊椎动物各纲中单配偶制也有其例。因为晚成鸟刚孵化的幼雏的发育很不完善,更需要双亲的共间抚育,所以多数晚成鸟是单配偶制,而早成鸟以多配偶制居多。例如北温带的许多鸣禽H食虫性小鸟。由于其作为食物资源的昆虫最丰富的时间是春季,有充分保证的期限也不很长,所以雏鸟必须迅速成长发育,并且缩短育雏期也能减少遭捕食危害的概率。待繁殖期一过,常集成较大的鸟群,过游荡的生活,不再在自己的领域上过“家庭”生活。
(2) 一雄多雌制。如前所述,一雄多雌制是动物圾常见的婚配制度,例如海狗科(OtariDAe)。海狗营集群生活,尤其在繁殖期内。通常雄兽首先到达繁殖基地,例如岩岸、沙滩或大块浮冰,随种而异,争夺并保护领域。雌兽到达较晚,不久就生胄幼仔。一只雄兽独占雌兽少则3只,多至40只以上,可以称为“闺房’’(harem)。闺房大小(指雌兽数)随种而异,也因雄兽凶猛程度和体力而不同。通常在产仔后不久,雄兽就在陆上与领域内的雌兽交配,怀孕期为250~365天。不少种类的海狗有孕妊潜伏期记载。
(3)一雌多雄制。与一雄多雌制相比较,动物中一雌多雄制是很稀见的,据统计,鸟类中只占1%。典型的例子是距翅水雉鸟。由于栖息地有限,种群中每年只有一部分雌鸟繁殖。雌鸟与若干只雄鸟交配,在不同地方产卵。与一雄多雌制相反,伏窝和育雏由雄鸟负担。两性异形也很明显,但雌鸟个体大于雄鸟,还更具进攻性,并协助雄鸟保护领域。有趣的是如果捕食者杀害了窝雏,雌鸟几乎能立即与雄鸟再次交配,几天后就产出一窝新卵。实际上距翅水雉的雌鸟已特化成为产卵“机器”,而在伏窝和育雏上,与一般的一雄多雌相比较,雌雄职责倒了过来。
(4)混配制。婚配方式不固定,配偶也不固定。鸟类中约有6%的种类是混配制的,哺乳动物如生活在苏格兰东北部的里氏田鼠灵长类中部分猩猩也是混配制的。还冇许多动物种群的婚配制度有可塑性,在不同的生长季或生活史的不同阶段,其婚配制度会发生变化。如北美的部分鸟类,在食物资源比较紧张的情况下,采用单配偶制;在资源很丰富的情况下,则采用一雄多雌制。
领域
领域(territory)是指一个动物或一个特定的动物群有选择地占领的,并加以守卫和防御,排斥同类个体或群体人侵的空间。而动物占领和保卫领域的行为则称为领域行为(territorialbehavior)。领域行为实际上起着减少同种动物之间战斗的作用,它对于控制种群密度、促进种群的稳定、对侵略性的抑制等都具有重要的现实意义。
领域主要特征
通常动物领域的主要特征体现 在如下几个方面:①领域是一个固定的空间区域,其大小可随时间和生态条件的不同而有所调整。几乎占有领域的边界都有明确的规定。②领域是受领域占领者积极保卫和防御的区域。领域多是通过争斗来划定的,范围一经确定,便各守一方,并通过不同的“标记行为”显示其所占有的领域,例如身体的姿态、运动、发出的叫声或产生出的特殊气味。鸟类通常从一棵树飞到另一棵树来巡视自己的领域。它们对领域边界外的对手毫不在意,但可向曾经越过边界但此刻已经远遁的对手发起攻击。虽然领域通常是由雄性动物建立的,但当雄性保卫巢室、孵卵及照料幼崽时,雌性便承担起守卫领域的任务。有时雄性和雌性动物只是防御同性别的对手。因此,最强的雄性和最强的雌性的组合就能占据最理想的领域。某些鸟如鹳和夜苍鹭,吸引它们的主要是筑巢地而不是配偶,如果前一年的配偶未曾返冋,它们可以接受新配偶。③领域的使用具有排他性,即它是被某一个或者某一些个体所独自占有的。通常,一个领域是由一个雄性动物建立起来的,其他雄性动物不容许进人其中,动物在它自己的领域内实际上是无敌的,这一点它的对手深信不疑。雄性动物在领域内接受雌性,进行求爱、建巢、抚育幼崽的活动,进食也通常在领域内进行。不能守卫领域的雄性动物不能进行繁殖。
领域大小规律
决定领域面积的基本规律如下:(1)领域面积随领域占有者的体重而扩大。因为领域大小必须以能保证供应足够的食物资源为前提,所以动物越大,需要资源越多,领域面积也就越大。(2) 食肉性种类的领域面积较同样体重的食草性种类大,并且体重越大,这种差别也越大。其原因是食肉动物获取食物更困难,需要消耗更多的能量,包括追击和捕杀。(3)领域行为和面积往往随生活史周期性变化,尤其随繁殖节律而变化。例如,鸟类一般在营巢期中领域行为表现最强烈,领域面积最大。
社会等级(Social hierarchy)是指种群中各个动物的地位具有一定顺序的等级现象。社会等级形成的基础是支配行为,或称支配-从属(dominant-submissive)关系。有关这方面研究得比较多的是鸡群中的啄击等级。当一群母鸡聚集在一起,并形成一个相对稳定的群体时,有一只母鸡对其他母鸡占有优势。它能啄击任何其他母鸡,而不会受到回啄。第二只母鸡也能啄击除第一只母鸡外的其他母鸡,第三只母鸡能啄击除前两只母鸡外的其他母鸡,如此类推,直到最后一只不幸的母鸡,它遭受所有母鸡的啄击,而不能冋啄,从而处于等级的最底层。在等级系统中位置较高的母鸡在占据食槽、进窝等方面具有特权,她们总是显得充满自信、风度翩翩、踌躇满志,而那些等级低下的母鸡看起来则是羽毛不整、情绪低沉、胆怯地在鸡群周围徘徊。有时,一个母鸡可能离开一个鸡群,加入另一鸡群,它们在不同的鸡群中可能有不同的等级。似在一个稳定的鸡群中,等级地位在一个相当长的时间内会保持不变。当一只新来的母鸡加入一个鸡群时,开始时她通常处于较低的等级,但经过实际的较量后,她会取得符合她实际情况的等级。当一个群体在建立优势等级时,须经历频繁而激烈的战斗,何等级一但建立,一仰首、一低头,就足以表示出优势或恭顺,因时群体中不再有战斗,生活和谐,井然有序。如果群体受到干扰(例如有一些新成员加入),那么等级系统就需要重建。公鸡一般不啄击母鸡,何他们也有自己的啄击等级。因此,在一个繁殖群体中,常常存在着两个等级系统。分属于不同的性别。占据优势的公鸡在生殖上是最成功的,而等级较低的公鸡,在生殖上机会较少,其至没有机会传播他们的基因。鸡没有稳定的配偶关系,有些动物则建立永久的配偶关系,这些动物同时也有等级系统。当一个雌性动物与一个雄性动物结成终身配偶时。她们就放弃原先在雌性中的等级地位,并取得她们配偶相应的等级地位。
社会等级制在动物界中相当普遍,包括许多鱼类、爬行类、鸟类和兽类。通过研究,已得到一些普遍的规律。高地位的优势个体通常较低地位的从属个体身休强壮、体重大、性成熟程度高,具有打斗经验。其生理基础是血液中存较高浓度的雄性激素(睾酮)。若给母鸡注射睾酮,她的等级地位会提高,若切去其卵巣,则其等级下降。猕猴群体社会中,占优势的雄猴体内睾酮的含量远高于战败的雄猴。
同物种之间,有时也存在优势等级。例如大兰苍鹭在各种苍鹭中占据高卨的优势等级。这种遗传决定的特性减少了不同物种之间的冲突。
任何动物都不是孤立地生活在自然界中的,它们总是组成一个小的生活群体。尽管有一些喜欢独来独往,但至少它们在交配时需要与异性接触。在接触过程中,它们的鸣叫,彼此间互相的触摸,甚至一些化学物质的释放,使得它们声息相通,行动一致,即动物之间的交流存在通信行为。所谓通信(communication)就是指个体通过释放一种或是几种刺激性信号,引起接受个体产生行为反应。根据信号的性质和接受的感官,可以把通信分为视觉的、化学的和听觉的等。信息传递的目的很广,如个体的识别,包括识别同种个体、同社群个体、同家族个体等,亲代和幼仔之间的通信,两性之间求偶,个体间表示威吓、顺从和妥协,相互警报、标记领域等。从进化观点而言,所选择的应为传递方便、节省能量消耗、误差小、信号发送者风险小、对生存必需的信号。世代之间的信号传递包括通过学习和通过遗传两类。通信是行为生态学研究的一个丰富而引人入胜的领域。
利他行为是另一种社会性相互作用。利他行为(altruism)是指一个个体牺牲自我而使社群整体或其他个体获得利益的行为。利他行为的例子很多,尤其是社会昆虫。白蚁的巢穴如被打开,工蚁和幼虫都向内移动,兵蚁则向外移动以围堵缺口,表现了“勇敢”的保卫群体的利他行为。工蜂在保卫蜂巢时放出毒刺,这实际上是一种“自杀行动“。亲代关怀(parental care)也是一种利他行为,亲代为此要消耗时间和能量,但能提高后代的存活率。一些鸟类当捕食者接近其鸟巢和幼鸟时佯装受伤,以吸引捕食者追击白己而将其引开鸟巢,然后自己再逃脱。不少啮齿类在天敌逼近时发出特有鸣叫声或以双足敲地,向周围的鼠发出报警信号,而发信号者反而更易引起捕食者的注意。
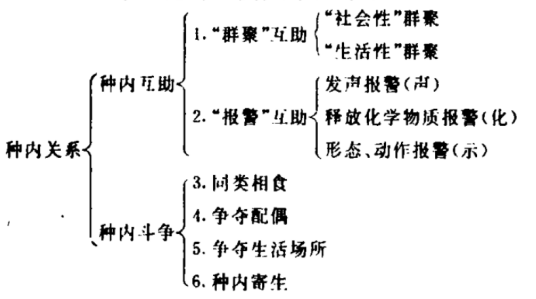
指同种生物个体之间,由于争夺食物、空间及配偶而发生斗争的现象。林地中的同种植物随种群密度的增大,对空间、养料、光照的争夺愈加剧烈,斗争的结果,优势个体与劣势个体之间生长的差距会愈加显著。
种内互助:是指同种生物个体之间的分工协作。这对于生物的生存和繁衍是有利的,如成群的狼通过分工合作,可以很容易地捕获到猎物。社会性昆虫更是将种内互助衍化到了分工明确的程度,如蚂蚁,工蚁专门负责采集食物、养育后代、修建巢穴;兵蚁具有强大的口器,专门负责保卫丁作;蚁后则成为专门产卵的生殖机器。还有鸿雁迁飞时,总是排着一列纵队或呈“人”字形雁阵,这种排列方法有助于鸿雁飞翔。因为飞在前面的大雁拍打一下翅膀,能使气流上升,后面的鸿雁就可借助这股气流飞翔,这样既能使雁群飞得更快,也能减少体力消耗。
下一篇 香藭散