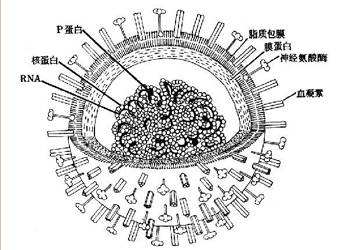
-
嵌合病毒 编辑
黄热病毒属(1qav/virus)包含80多个成员,属于正链RNA病毒,其中半数以上的病毒能够诱发人和动物产生疾病,患病率高而治愈率低,容易留下一些严重的后遗症,给人类健康带来很大威胁,如流行性乙型脑炎病毒(JEV)、登革热病病毒(DEN)、黄热病毒(YFV)、蜱传脑炎病毒(TBEV)等。一直以来人们对RNA病毒的了解都比较缺乏,无论是分子水平上的机理研究,还是宏观上疫苗的研制、药物的开发,其进展都非常缓慢
 嵌合病毒
嵌合病毒
嵌合病毒是一种基因工程重组病毒,在制备技术上并不存在太大问题,但作为重组病毒,在安全性上还有一定的风险,在自然条件下,有病毒之间自发产生重组的例子,可能获得一些意想不到的新型病毒,比母体株毒力更强,对生物体更具有威胁性。所以在制备嵌合病毒时,要采用已知特性的病毒株作为嵌合病毒的骨架结构,特别是多选用减毒株作为载体病毒黄热病毒的嵌合病毒采用的载体主要有以下几种。
YF一17DD 疫苗株 YF一17DD是预防黄热病毒感染的一株减毒活疫苗】,始用于1936年,在过去的6O多年中,有超过3亿人接种过该疫苗,安全有效。该疫苗株免疫原性很好,单剂量接种后10d内,近lo0%的接种者产生中和抗体,免疫力持久甚至能够维持终身。将YF一17DD疫苗株和原病毒株YF—Asibi进行序列比较后发现,该病毒的抗原表位多集中在E基因,构建嵌合病毒多以这一段基因作为靶基因。这样构建的嵌合病毒中,由YF一17DD株保留的NS蛋白基因只是诱发针对该蛋白的CTLs和非中和性抗体,而不具有交叉保护性,所以嵌合病毒作为疫苗使用时不会引起一般重组病毒带来的抗载体免疫反应,即使曾经接种过YF一17DD的接痘者再接种嵌合病毒也是安全有效的,可以作为一个理想的活疫苗株嵌合病毒载体。1989年Rice等151构建了YF一17DD的感染性克隆,为构建嵌合病毒提供了方便,文献中报道的有关黄热病毒嵌合株多以此病毒株作为载体病毒。已经成功构建了包括JEV、TBE、WNV、DEN等4个血清型病毒的嵌合体。
YF一17DD 疫苗株
DEN一4变异株 登革病毒分为4个血清型,这4型病毒结构蛋白氨基酸序列同源性高达62%-74%,并且它们之间还有很好的交叉免疫原性,一直以来,人们试图构建登革病毒的4价疫苗,Bray等从1991年开始登革病毒嵌合病毒的研究,以DEN一4814669株全长克隆作为载体,构建了包括DEN—l到DEN一3的嵌合病毒,结果发现几株病毒的毒性并没有太大改变,他们认为若能够以一个毒性低的DEN-4病毒作为载体骨架,可能会使嵌合病毒毒性下降,由于3端和5端非编码区在病毒的复制中可能也起到调控作用,他们在构建的DEN一4感染性克隆的基础上,删除其3端和5端非编码区的部分序列,结果筛选到一株减毒株,其3端去掉了3O个核苷酸,毒性降低但是免疫原性并没有改变。用这个减毒株作为载体和其它3个血清型的登革病毒构成嵌合病毒后,果然有效地降低了病毒毒性;其后这个载体也用于构建黄热病毒属其他成员的嵌合病毒。
DEN一4变异株
DEN一2 PDK一53减毒株 该减毒株也主要是用于合成登革病毒的嵌合疫苗,2O世纪8O年代初期,泰国Mahidol实验室将4种血清型的登革病毒经过在细胞传代中后获得各自的减毒株,其中以DEN一2PDK一53减毒株减毒效果最好,免疫保护力最强最持久。将该减毒株和野生株进行比较,发现核苷酸水平上的变异主要在非结构区,说明该病毒株和毒力有关的基因可能在非结构区,而黄热病毒属其他成员的毒力基因多位于E蛋白,这样构建的嵌合病毒减毒机制主要由供体株的毒性起主要因素,而由DEN一2PDK-53减毒株构建的嵌合疫苗毒性的减弱由供体和受体共同决定,更有利于减毒株的获得
DEN一2 PDK一53减毒株
DEN一2减毒株作为嵌合病毒的载体还有另外一个好处,登革病毒4个血清型之间产生交叉免疫力,所以异型病毒的二次感染由于抗体依赖的增强作用可以导致登革出血热/登革热休克综合征(DHF/DSS)的发生,理想的登革4价疫苗诱导的免疫力要针对4种血清型病毒,有效而且要持久,其中任何一种血清型抗体的消减都有发生DHF/DSS的可能。流行病学资料表明,DEN几个血清型感染人的概率是不同的,其中以DEN一2的感染概率最高,所以由其它几种血清型病毒诱发的免疫反应很易识别DEN一2病毒,由此DEN一2病毒诱发DHF/DSS的概率明显大于DEN—l、一3、一4;另外DEN一2病毒诱导的特异免疫力又明显弱于其它3型,所以制备以DEN一2为基础的4价疫苗和传统疫苗比较,减少了DEN—l、一3、一4的4种血清型NS3蛋白诱发的交叉免疫反应之间的干扰作用,减少了DHF/DSS的可能,同时又能有效而持久地诱导机体产生针对4种血清型的免疫反应,从而起到预防作用。
克隆法 嵌合病毒的获得大多是利用感染性克隆技术,已有部分黄热病毒属病毒获得了感染性克隆,如YFV、TBEV、Kunjin病毒、墨累河谷脑炎病毒(MVE)、西尼罗河病毒(WNV)、DEN4型病毒,这些克隆包含有病毒的全部基因,能够在宿主菌中稳定地复制和传代,以作为种子批保存。当构建嵌合体时,在供体基因和受体基因的连接处,通过同义突变或者保守突变,为受体株和供体株基因组添加合适的酶切位点,然后将两者利用酶切位点连接起来,制备嵌合性克隆,体外经RNA聚合酶作用转录出嵌合性转录子,转染细胞后重新获得嵌合病毒。这项技术的一个优点在于在它是在cDNA水平上的操作,突变率低、表型稳定,不像传统疫苗研制过程中容易发生一系列的生物特性改变。
克隆法
融合PCR 利用PCR技术制备嵌合病毒最大的优点在于方便快捷,过去几个月的工作几天就能完成。生物技术产品日臻完善,不少公司推出高保真、热稳定性好的逆转录酶和DNA聚合酶,如SuperscriptII逆转录酶、LAro~酶、Exraq酶、ExpandlongtemplatePCR聚合酶系统等,利用这些酶可以有效地扩增出病毒基因组大片段(>5kh)甚至全长的cDNA。用4条引物、3次PCR可以把两个异质基因片段连接在一起。另有报道利用3个引物、两次PCR即可制备嵌合基因,如YF—MOD嵌合病毒。
融合PCR
与克隆技术相比较,PCR方法更为灵活,可以选择任意位置进行嵌合;但是PCR具有一定的突变率,可能会带来一些致死性突变而使转录体不具有感染性,得不到恢复病毒从而前功尽弃,所以大多数实验还是采取克隆法。
灭活病毒之间的基因重组 是将两种或者两种以上的灭活病毒,在同一细胞内培养后,它们之间发生基因重组,有时会产生感染性病毒,这种方法又称为多重复合,可能由于灭活病毒受损的基因经重组后得以替补。如为了解包膜病毒的融合活性,利用黄热病毒YF一17DD和流感病毒X31两者不同的融合条件,以及核衣壳不同的形态,将YF一17DD通过酸处理失去融合活性,X31通过热处理灭活,两者在YF一17DD灭活的同一pH值下发生融和反应,只有发生融和反应的嵌合病毒具有感染性;流感病毒提供功能型膜蛋白融合成分,YF一17DD提供功能基因组,结果两个灭活病毒又重新构建成新的病毒。这项技术随机性很大,但是可能得到意想不到的结果,可以用于病毒功能的研究。
灭活病毒之间的基因重组
黄热病毒嵌合病毒的构建前提在于,所有的黄热病毒的病毒基因组的结构是高度保守的,它们均为单股正链RNA病毒,基因组全长约l1kb,5端有帽子结构,3端无poly(A)尾,一个长的开放阅读框架覆盖了大部分基因组,共编码3个结构蛋白和7个非结构蛋白,其中以E蛋白和免疫原性关系最为密切,所以构建的嵌合病毒一般都要取供体病毒的E基因;但并不是所有构建的嵌合性cDNA克隆都具有感染性,其主要影响因素如下。
技术方面 虽然工具酶的保真性和效率越来越好,但是黄热病毒长10kb以上,在基因操作中不可避免有突变的发生,而有些突变是致死性突变,使得构建的全长cDNA克隆或者嵌合体克隆没有感染性。另外这些突变还影响恢复病毒的生物学特性,如温度敏感性、噬斑大小、复制能力、嗜细胞能力都可能发生改变。
技术方面
嵌合位点 结构蛋白基因之间的连接处有一段亲脂区序列,是构建嵌合病毒时将两段异质核苷酸序列进行连接的嵌合位点处,这一段序列的保守性对于嵌合病毒的复制能力至关重要。如在prM和C基因之间有信号肽酶切割位点以及蛋白酶切割位点,Cau—four等在构建YF17D—DEN2嵌合病毒时选择这两处作为嵌合位点,结果在信号肽酶切割位点处嵌合的病毒有感染性,而在蛋白酶切割位点处进行连接无法获得病毒,因为在两个位点之间有一段亲脂性的转膜区,YFv这一区域比DEN一2该区域要多6个氨基酸;在蛋白酶切割位点处构建的嵌合体,由于这一转膜区缺少了6个氨基酸,蛋白酶和信号肽酶不能正确识别切割位点,从而影响prM蛋白和E蛋白的成熟,病毒的复制过程受到阻碍,病毒颗粒没法进行正常的包装,所以无法分泌出病毒颗粒。
嵌合位点
供体病毒基因 构建的嵌合病毒多为供体株提供P和E基因,人们也尝试过替代C基因或者部分基因,但多数没有成功,其可能的原因是位于C蛋白区域的保守序列与受体株病毒顺式结构不相容性,进而对嵌合病毒RNA的复制产生了毒性,如Kuniin病毒RNA复制必须有同源C蛋白上的第2位和第20位氨基酸的存,而异质基因的C蛋白不能协助Kunjin病毒复制,所以无法长成病毒颗粒;另外也可能是异质病毒之间的蛋白酶识别对方序列的能力低下,对于C—prM部位的切割效率低,影响病毒正常的装配和释放能力。
供体病毒
受体病毒基因 作为嵌合病毒载体,嵌合病毒中的大部分基因由受体株提供,嵌合病毒的复制特性也主要由受体株的基因控制,其中有些序列对于嵌合体的感染性有决定性作用,在构建嵌合体时,如果这部分序列受到破坏,就不能够获得恢复病毒。如在构建DEN一4和WNV的嵌合病毒时四,由WNVNY99的prM和E基因代替DEN一4相应位置的基因,在构建的18个克隆中仅有两个具有感染性,序列比较发现这两个克隆在prM和C蛋白之间的NS2A—NS2B蛋白酶切割位点下游,在转膜信号区域中都包含一基序,这一基序是酶切割位点下游的第3位及第6位分别为天冬氨酸和苏氨酸,另外16个克隆则没有该位点,实验表明这两个位点似乎和嵌合性克隆的感染性有关。
受体病毒
嵌合病毒的一个最大特点是表型多发生变化,如复制能力、毒力、抗原性等,可能的原因是由于缺乏校正功能的聚合酶存在,而有异质基因的正链RNA病毒,在病毒复制中具有较高的基因突变率,并且这些突变为了适应不同的宿主细胞,可能会发生一些特性改变,从而出现一些预想不到的结果。
生长特性改变 嵌合病毒的两个母体病毒的易感细胞不同,得到的嵌合病毒在不同细胞中复制时,由于缺乏有效的复制酶而引起生长特性的改变,其嗜细胞性也会有所改变,另外噬斑试验中噬斑形态
生长特性改变
大小都多有变化。如Chimeri—JE嵌合体在Vero、LLC—MK,、C6/36细胞中复制形式类似,而在MRC一5、FRhl细胞中虽然病毒滴度能够达到6log。0pfu/mL,但是却不能产生CPE,这些特性和母体株YFV以及JEV都截然不同。LGT1和E5是TBEV的两个弱毒株,与DEN一4病毒组成的嵌合体在猿猴细胞中复制能力下降,而转染蚊虫细胞复制能力不受影响,用在蚊虫细胞传代的嵌合病毒在猿猴细胞LLC—MK2中不能够形成噬斑,而在早期构建的DEN一4和另一个TBEV病毒株构建的嵌合体在猿猴细胞中却能够得到比DEN一4病毒滴度高1000倍的嵌合病毒。
神经内毒性和神经外毒性 神经内(外)毒性低而免疫原性好是一个理想疫苗应具备的品质,许多实验室经过多年研究虽然筛检出一些减毒株,但是两者多不能达到有效的统一。嵌合病毒技术以“取长补短”的原则,将两种病毒的免疫原性和毒性结合起来,以期研制理想疫苗,就得到的一些嵌合株来说,确实取得了一些令人满意的结果,如DEN一4和WNV的嵌合病毒与WNV相比高度减毒四,脑内接种戒奶小鼠,神经内毒性降低至1/28500;腹腔内接种小鼠,神经外毒性下降至1/10000,免疫原性好,能安全保护小鼠接受野生株攻击。另外如ChimeriVax—JE嵌合病毒毒性也较母体株YFV和JEV呈大幅度下降,而单剂量又具备很好的免疫保护力;还有DEN—TBEV、YFV-DEN、YFV—WNV等嵌合病毒株的毒性都有所下降,这些嵌合体都在进一步的研究中。
神经内毒性和神经外毒性
基因稳定性 因为RNA病毒易以准种的形式出现和易变异的特性,作为活疫苗来使用,一个重要的条件是基因型要稳定,这一点是疫苗安全性的重要表现。嵌合病毒的稳定性主要通过返祖实验检定,即在乳鼠脑内和细胞中进行培养,观察有无明显表型变化,必要时进行核苷酸鉴定,如ChimeriVax-JE嵌合病毒115.161在细胞内传代l8次以及在乳鼠脑内传代6次并没有毒力的回升,对减毒位点进行测序也没有发生核苷酸改变。而有的嵌合病毒稳定性不够好如DEN一2/DEN一4、DEN一2/DEN—l复制能力较弱,而经过几次传代后得到较高的病毒滴度,是因为发生了一些突变。在体外转录时产生了转录体的杂和体,在转染后经过细胞复制,复制能力增强的病毒逐渐占主要优势,在LLC—MK2细胞中病毒传代可以保持较高的遗传稳定性,所以在制备嵌合疫苗时,应尽量减少病毒在哺乳动物细胞中传代的次数。
基因稳定性
在黄热病毒嵌合病毒发展的这十几年中,人们尝试构建了多种嵌合病毒,其中大部分是以制备疫苗为目的,另外还用于病毒的分子生物学特性研究。
分子生物学特性研究 型内嵌合病毒可以用于了解病毒复制能力,进一步了解各基因的功能。如构建同型不同株之间DEN一216681/PDK一53、DEN25ol/04株之间的型内嵌合疫苗,研究DEN一2型病毒结构蛋白区对乳鼠神经毒性的影响。可以用于推测病毒毒力位点,由2个JEV病毒株cDNA克隆提供5端,3个病毒株cDNA提供的3端进行组合,感染细胞后重新获得6种重组JEV病毒。根据噬菌斑大小筛选到3个减毒株,比较6种重组病毒株和野生株的基因序列,发现仅仅是E138(Glu—Lys)一个氨基酸位点不同,说明这个位点和毒力之间关系密切;另外发现该突变株虽然毒性降低,但是脑内注射接种小鼠后仍能在脑内复制,但是病毒滴度比较低,看来其减毒机制不在于病毒的复制,而可能是影响复制的早期机制,在细胞之间传递速度比较慢,从而使免疫系统能及时清除病毒。
分子生物学特性研究
型间嵌合病毒也可以用于分子生物学特性的研究,如黄热病毒和日本脑炎病毒虽然同属黄热病毒属,但是有很多生物学特性都不尽相同,如抗原性相差很远、中间宿主不同、引起人体发病的症状也截然不同;两者虽然从同一病毒得来,但是在进化的过程中发生了很大的变化,构建的嵌合病毒YF/JE(S)其生物学特性也有所改变,如神经内毒性和神经外毒性都有所降低,和其供体株SAl4一l4—2相似;而构建的另一个嵌合株YF/JE(N)神经毒性却很高,和供体株Nakayama特性一致;这说明确实如原先预料,和减毒有关的表位主要存在E蛋白中,而同时也发现,和原来的SA14-14-2相比较,嵌合株在细胞中复制能力升高,病毒滴度较高,免疫原性增加,这又说明是YF一17DD病毒的非结构蛋白发挥作用。
候选疫苗 ChimeriVax-JE嵌合病毒
候选疫苗
日本脑炎是一种影响中枢神经系统的疾病,主要流行于亚洲,但是近年在澳洲、美洲也有病例发生,每年官方报道全球有5万病例,其中有l万例是致死性的;虽然有活疫苗、灭活疫苗的使用,但是这些疫苗具有免疫原性不够、价格偏高、制作工艺不被WHO认可等劣势,所以还有必要开发研制新疫苗。1999年Chambers等以SA14-14-2的prM和E基因代替YF一17DD疫苗株的相应基因构建了嵌合病毒ChimeriVax—JE[15,16,,该病毒能在WHO认可的细胞中很好地被培养,恢复病毒和YF一17DD、SA14—14-2相比较,以6log0pfu的嵌合病毒脑内接种3-4周龄小鼠无一例发病,而接种YF-17DD的小鼠全部死亡;另外用3.2log0pfu剂量的SA14-14-2接种,能在85%一95%的接种者中产生中和抗体,抗体平均滴度能够达到PRNT~o<30,血清阳转率小于80%;而接种4~51og0pfu的嵌合疫苗后,能够达到100%的血清阳转率,接种后30dPRNT能够达到129~327,这些结果表明构建的嵌合病毒神经内毒性呈大幅度降低;用乳鼠以及猴子进行安全试验检测,进一步证实了该病毒的弱神经毒性,内脏毒性也下降。该疫苗已经通过一期临床试验,是很有前途的疫苗候选株。
登革病毒多价疫苗
登革病毒是诱发疾病最多的黄热病毒,在人口聚集的热带及亚热带时有爆发,因为登革病毒4个血清型之间可以产生交叉保护力,所以感染一种病毒后,能够对另外3种血清型病毒的抗体反应。在机体遭受另一种血清型病毒的感染后,由于抗体依赖的复制能力增强作用可能诱发DHF/DSS的发生,因此理想的疫苗应该能够产生针对4个型病毒的中和抗体和T细胞免疫。主要有两个实验室分别以DEN一2PDK一53和YF一17DD作为受体株构建了包括DEN一1到DEN-4型的嵌合株,结果有很好的免疫原性、较高的复制效率和稳定性,在动物实验中证明毒性减低,小鼠效力实验安全有效,有望成为登革病毒疫苗的发展方向。
另外还有YF/TBE、YF/WNV、DEN4/TBE的报道,都出现一些令人满意的特性,进一步研究正在进行中。嵌合病毒的出现是获得减毒活疫苗的一个新途径,和传统方法相比具有目标明确、免疫效果好、减毒稳定的优势,但该技术还处于探索阶段,有时结果很出人意料,所以在构建一个新的嵌合体时,要时刻注意安全性问题,否则会适得其反,创造出新的毒性更强的病毒,为人类带来危害。